 |
|
| No | Structure | COMMON NAME | NAME | DATA No | INFORMANT | SYMBOL | FORMULA | MOL.WT(ave) | Download | BIOOGICAL ACTIVITY | PHYSICAL AND CHEMICAL PROPERTIES | SPECTRAL DATA | CHROMATOGRAM DATA | SOURCE | CHEMICAL SYNTHESIS | METABOLISM | GENETIC INFORMATION | NOTE | REFERENCES | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MELTING POINT | BOILING POINT | DENSITY | REFRACTIVE INDEX | OPTICAL ROTATION | SOLUBILITY | UV SPECTRA | IR SPECTRA | NMR SPECTRA | MASS SPECTRA | OTHER SPECTRA | ||||||||||||||||||
| 1 | 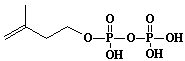 |
Isopentenyl diphosphate |
3-Methyl-3-butenyl diphosphate |
IIP0001 | Hiroshi Sagami |
IPP |
C5H8O7P2 | 242.060 | The compound is an active isoprene unit commonly occurring in various organisms. |
Procedures for the synthesis and purification of this compound, other 20 isoprenoid diphosphates and methanediphosphate analogues from the corresponding alcohols are found in (Ref. 0014). The synthesis of isoprenoid (phosphinylmethyl)phosphonates are described in (Ref. 0015) . Negative ion fast atom bombardment mass spectrometry (FABMS) has been used to obtain mass spectra of the naturally occurring isoprenoid metabolites including isopentenyl diphosphate, dimethylallyl diphosphate, geranyl diphosphate, and farnesyl diphosphate, presqualene diphosphate, and of several synthetic analogs including methylene-bridged diphosphates, bisphosphates, and nitrogen-containing analogs.(Ref. 0023) The study of plant polyisoprenoid alcohols including polyprenols and dolichols has been reviewed in (Ref. 0024). |
||||||||||||||||||
| 2 | 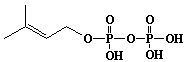 |
Dimethylallyl diphosphate |
3-Methyl-2-butenyl diphosphate |
IIP0002 | Hiroshi Sagami |
DMAPP |
C5H8O7P2 | 242.060 | This compound can be synthesized from dimethylallyl alcohol (Ref. 0014). |
The steroisomer of isopentenyl diphosphate and the product formed by the action of isopentenyl diphosphate isomerase. |
The nucleotide sequence of isopentenyl diphosphate:dimethylallyl diphosphate isomerase (Saccharomyces cerevisiae) has been submitted to the GenBankTM/EMBL data bank with accession number J05090.(Ref. 0010) |
|||||||||||||||||
| 3 | 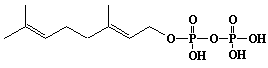 |
Geranyl diphosphate, Geranyl pyrophosphate |
[2E]-3,7-Dimethyl-2,6-octadien-1-yl diphosphate |
IIP0003 | Yoshichika Yoshioka |
GOPP, GPP |
C10H20O7P2 | 314.209 | Metabolic labeling of rat liver slices with mevalonic acid revealed the accumulation of E,E,E-geranylgeranyl (di)phosphate as well as dolichyl (di)phosphate (C85 and C90) and dehydrodolichol (C85 and C90), but no accumulation of Z,E,E-geranylgeranyl (di)phosphate or E,E-farnesyl (di)phosphate was detected(Ref. 0026). |
The compound can be synthesized from geraniol.(Ref. 0014) |
The product formed by the action of geranyl diphosphate synthase and the first intermediate product by the action of farnesyl diphosphate synthase from dimethylallyl diphosphate and isopentenyl diphosphate. The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capacity was as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and dolichol were not inhibitors(Ref. 0039). |
|||||||||||||||||
| 4 | 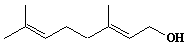 |
Geraniol |
[2E]-3,7-Dimethyl-2,6-octadien-1-ol |
IIP0004 | Yoshichika Yoshioka |
GOH |
C10H18O | 154.249 | The 2H-labeled geraniol gave rise to 2H NMR powder patterns interpretable in terms of quadrupole splittings and splin-lattice relaxation times. Spin-lattice relaxation time measurements revealed high rates of motion for geraniol relative to cholesterol in similar membrane hosts and revealed correlation times close to the fatty acyl methyl termini in phosphatidylcholine.(Ref. 0038) |
The product derived from geranyl diphosphate. The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capasity were as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and dolichol were not inhibitors(Ref. 0039). |
||||||||||||||||||
| 5 | 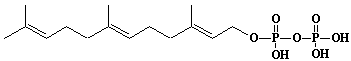 |
Farnesyl diphosphate, Farnesyl pyrophosphate |
[2E,6E]-3,7,11-Trimethyl-2,6,10-dodecatrien-1-yl diphosphate |
IIP0005 | Yoshichika Yoshioka |
FPP, FOPP |
C15H28O7P2 | 382.326 | Rat liver microsomes catalyzed the formation of Z,E,E-geranylgeranyl diphosphate from farnesyl diphosphate and isopentenyl diphosphate in the presence of Triton X-100(Ref. 0026). Metabolic labeling of rat liver slices with mevalonic acid revealed the accumulation of E,E,E-geranylgeranyl (di)phosphate as well as dolichyl (di)phosphate (C85 and C90) and dehydrodolichol (C85 and C90), but no accumulation of Z,E,E-geranylgeranyl (di)phosphate or E,E-farnesyl (di)phosphate was detected(Ref. 0026). HMG-CoA reductase inhibitors induce apoptosis in mouse proximal tubular cells in primary culture. Geranylgeranylpyrophosphate and mevalonate completely reversed the effect of the inhibitors, while farnesylpyrophosphate partially reversed it and dolichol had no effect(Ref. 0064). |
The compound can be synthesized from farnesol.(Ref. 0014) |
The rate of de novo synthesis of geranylgeranyl diphosphate from mevalonic acid is comparable to that of farnesyl diphosphate(Ref. 0078). Geranylgeranyl diphosphate synthase catalyzing the single condensation between isopentenyl diphosphate and farnesyl diphosphate. The enzyme required Mg2+ and Mn2+ for maximum activity. Octylglucoside showed a stimulatory effect on the enzyme activity. Farnesyl diphosphate is the common intermediate at the branch point for the synthesis of geranylgeranylated proteins as well as cholesterol, ubiquinone, dolichol, and farnesylated proteins(Ref. 0070). The uptake of F-PP and GG-PP was studied using primary cultures of chromaffin cells. Dolichylmonophosphate and isopentenylpyrophosphate promoted the uptake(Ref. 0079). The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capasity were as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and dolichol were not inhibitors(Ref. 0039). |
|||||||||||||||||
| 6 | 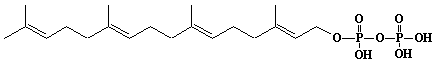 |
Geranylgeranyl diphosphate, Geranylgeranyl pyrophosphate |
[2E,6E,10E]-3,7,11,15-Tetramethyl-2,6,10,14-hexadecatetraen-1-yl diphosphate |
IIP0006 | Yoshichika Yoshioka |
GGOPP, GGPP |
C20H36O7P2 | 450.443 | Rat liver microsomes catalyzed the formation of Z,E,E-geranylgeranyl diphosphate from farnesyl diphosphate and isopentenyl diphosphate in the presence of Triton X-100(Ref. 0026). HMG-CoA reductase inhibitors induce apoptosis in mouse proximal tubular cells in primary culture. Geranylgeranylpyrophosphate and mevalonate completely reversed the effect of the inhibitors, while farnesylpyrophosphate partially reversed it and dolichol had no effect(Ref. 0064). |
The compound can be synthesized from geranylgeraniol .(Ref. 0014) |
The product formed by the action of geranylgeranyl diphosphate synthase. The rate of de novo synthesis of geranylgeranyl diphosphate from mevalonic acid is comparable to that of farnesyl diphosphate(Ref. 0078). Geranylgeranyl diphosphate synthase catalyzing the single condensation between isopentenyl diphosphate and farnesyl diphosphate. The enzyme required Mg2+ and Mn2+ for maximum activity. Octylglucoside showed a stimulatory effect on the enzyme activity. Farnesyl diphosphate is the common intermediate at the branch point for the synthesis of geranylgeranylated proteins as well as cholesterol, ubiquinone, dolichol, and farnesylated proteins(Ref. 0070). Purification and properties of geranylgeranyl-diphosphate synthase from bovine brain(Ref. 0080). The uptake of F-PP and GG-PP was studied using primary cultures of chromaffin cells. Dolichylmonophosphate and isopentenylpyrophosphate promoted the uptake(Ref. 0079). The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capasity were as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and dolichol were not inhibitors(Ref. 0039). |
The nucleotide sequences of yeast Saccharomyces cerevisiae and human geranylgeranyl diphosphate synthase have been reported in (Ref. 0011)and (Ref. 0016). African swine fever virus trans-polyprenyltransferase also catalyzes mainly geranylgeranyl diphosphate from farnesyl diphosphate and isopentenyl diphosphate. (Ref. 0013) |
||||||||||||||||
| 7 | 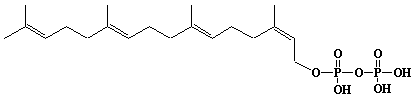 |
w-E,E,Z-Geranylgeranyl diphosphate, Cis-Geranylgeranyl diphosphate, geranylneryl diphosphate |
[2Z,6E,10E]-3,7,11,15-Tetramethyl-2,6,10,14-hexadecatetraen-1-yl diphosphate |
IIP0007 | Hiroshi Sagami |
C20H36O7P2 | 450.443 | The compound can be synthesized from cis-geranylgeraniol .(Ref. 0014) |
The product formed by the action of cis-geranylgeranyl diphosphate synthase and the first product by the action of dehydrodolichul diphosphate synthase from farnesyl diphosphate and isopentenyl diphosphate.(Ref. 0026) |
|||||||||||||||||||
| 8 | 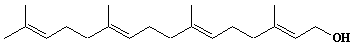 |
Geranylgeraniol |
[2E,6E,10E]-3,7,11,15-Tetramethyl-2,6,10,14-hexadecatetraen-1-ol |
IIP0008 | Yoshichika Yoshioka |
GGOH |
C20H34O | 290.483 | Geranylgeraniol had a potent apoptosis-inducing activity in HL-60 cells.(Ref. 0040) Geranylgeraniol causes a decrease in levels of calreticulin and tyrosine phosphorylation of a 36-kDa protein prior to the appearance of apoptotic features in HL-60 cells.(Ref. 0041) Geranylgeraniol potently induces caspase-3-like activity during apoptosis in human leukemia U937 cells.(Ref. 0042) |
The product derived from geranylgeranyl diphosphate. |
||||||||||||||||||
| 9 | 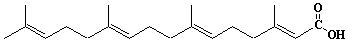 |
Geranylgeranic acid, geranylgeranoic acid |
[2E,6E,10E]-3,7,11,15-Tetramethyl-2,6,10,14-hexadecatetraenic acid |
IIP0009 | Yoshichika Yoshioka |
GGA |
C20H32O2 | 304.467 | Synthetic 4,5-didehydro geranylgeranoic acid, a potent ligand both for cellular retionoic acid-binding protein and for nuclear retinoid receptors, induced apoptosis in human hepatoma-derived cell line HuH-7 but not in primary hepatocytes, although all-trans or 9-cis retinoic acid did not induce any growth inhibition(Ref. 0081). A synthetic geranylgeranoic acid (GGA) induced apoptotic cell deth in a human hepatoma cell line, HuH-7, but not in mouse primary cultured hepatocytes. Prior to chromatin condensation, GGA induced a dramatic loss of the mitochondrial membrane potential in 1 hour and in a dose dependent manner in HuH-7 cells, but not in the primary hepatocytes. GGA induces apoptosis in hepatoma cells through derangement of mitochondrial function and subsequent activation of the cysteine protease cascade(Ref. 0082). |
The product derived from geranylgeranyl diphosphate. |
||||||||||||||||||
| 10 | 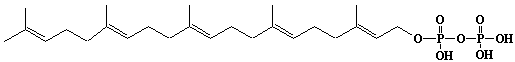 |
Geranylfarnesyl diphosphate |
[2E,6E,10E,14E]-3,7,11,15,19-Pentamethyl-2,6,10,14,18-eicosapentaen-1-yl diphosphate |
IIP0010 | Hiroshi Sagami |
GFPP |
C25H44O7P2 | 518.560 | The product formed by the action of geranylfarnesyl diphosphate synthase. |
|||||||||||||||||||
| 11 | 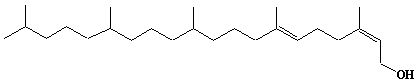 |
Pentaprenol, Prenol-25 |
[2Z,6E]-3,7,11,15,19-Pentamethyl-2,6-eicosadien-1-ol |
IIP0011 | Hiroshi Sagami |
C25H48O | 364.648 | The function is not known. |
The compound was isolated from the leaves of potato, Solanum tuberosum. (Ref. 0001) and comprises a phytyl residue and a Z-a terminal residue. |
|||||||||||||||||||
| 12 | 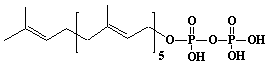 |
Hexaprenyl diphosphate, w-E,E,E,E,E-Hexaprenyl diphosphate, trans-Hexaprenyl diphosphate |
[2E,6E,10E,14E,18E]-3,7,11,15,19,23-Hexamethyl-2,6,10,14,18,22-tetracosahexaen-1-yl diphosphate |
IIP0012 | Hiroshi Sagami |
C30H52O7P2 | 586.677 | The product formed by the action of hexaprenyl diphosphate synthase. |
||||||||||||||||||||
| 13 | 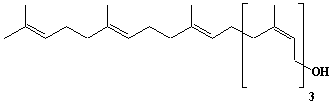 |
Betulaprenol-6, Hexaprenol |
[2Z,6Z,10Z,14E,18E]-3,7,11,15,19,23-Hexamethyl-2,6,10,14,18,22-tetracosahexaen-1-ol |
IIP0013 | Hiroshi Sagami |
C30H50O | 426.717 | |||||||||||||||||||||
| 14 | 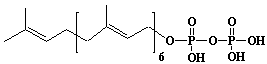 |
w-E,E,E,E,E,E-Heptaprenyl diphosphate, Heptaprenyl diphosphate |
[2E,6E,10E,14E,18E,22E]-3,7,11,15,19,23,27-Heptamethyl-2,6,10,14,18,22,26-octacosaheptaen-1-yl diphosphate |
IIP0014 | Hiroshi Sagami |
C35H60O7P2 | 654.794 | The product formed by the action of heptaprenyl diphosphate synthase. Isoprene double bonds are trans except for w-isoprene double bond. |
||||||||||||||||||||
| 15 | 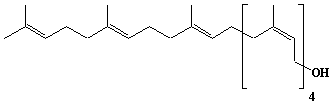 |
Betulaprenol-7, cis-Heptaprenol |
[2Z,6Z,10Z,14Z,18E,22E]-3,7,11,15,19,23,27-Heptamethyl-2,6,10,14,18,22,26-octacosaheptaen-1-ol |
IIP0015 | Hiroshi Sagami |
C35H58O | 494.834 | |||||||||||||||||||||
| 16 | 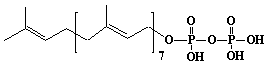 |
Octaprenyl diphosphate, trans-Octaprenyl diphosphate |
[2E,6E,10E,14E,18E,22E,26E]-3,7,11,15,19,23,27,31-Octamethyl-2,6,10,14,18,22,26,30-dotriacontaoctaen-1-yl diphosphate |
IIP0016 | Hiroshi Sagami |
C40H68O7P2 | 722.911 | The product formed by the action of octaprenyl diphosphate synthase. This compound contains trans isoprene double bonds except for w isoprene double bond. |
The amino acid sequence of Escherichia coli octaprenyl diphosphate synthase has (GenBank accession no. U18997) been reported in (Ref. 0018). |
|||||||||||||||||||
| 17 | 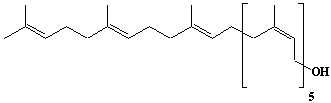 |
Betulaprenol-8, Octaprenol |
[2Z,6Z,10Z,14Z,18Z,22E,26E]-3,7,11,15,19,23,27,31-Octamethyl-2,6,10,14,18,22,26,30-dotriacontaoctaen-1-ol |
IIP0017 | Hiroshi Sagami |
C40H66O | 562.951 | |||||||||||||||||||||
| 18 | 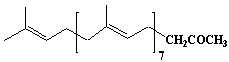 |
Bombiprenone |
[all-E]- 6,10,14,18,22,26,30,34-octamethyl-5,9,13,17,21,25,29,33-pentatriacontaoctaen-2-one |
IIP0018 | Hiroshi Sagami |
C43H70O | 603.015 | The compound was isolated from silkworm feces.(Ref. 0004) |
||||||||||||||||||||
| 19 | 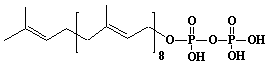 |
Solanesyl diphosphate, Nonaprenyl diphosphate |
[2E,6E,10E,14E,18E,22E,26E,30E]-3,7,11,15,19,23,27,31,35-Nonamethyl-2,6,10,14,18,22,26,30,34-hexatriacontanonaen-1-yl diphosphate |
IIP0019 | Hiroshi Sagami |
C45H76O7P2 | 791.028 | The product formed by the action of nonaprenyl diphosphate synthase. |
The amino acid sequence of solanesyl diphosphate synthase from Rhodobacter capsulatus (Ref. 0012). |
|||||||||||||||||||
| 20 | 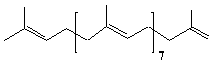 |
Nor-solanesene |
[all-E]- 2,6,10,14,18,22,26,30,34-Nonamethyl-1,5,9,13,17,21,25,29,33-pentatriacotanonaene |
IIP0020 | Hiroshi Sagami |
C44H72 | 601.042 | The compound was isolated from tabacco.(Ref. 0005) |
||||||||||||||||||||
| 21 | 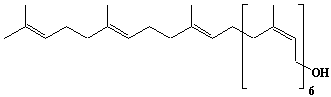 |
Betulaprenol-9, Nonaprenol |
[2Z,6Z,10Z,14Z,18Z,22Z,26E,30E]-3,7,11,15,19,23,27,31,35-Nonamethyl-2,6,10,14,18,22,26,30,34-hexatriacontanonaen-1-ol |
IIP0021 | Hiroshi Sagami |
C45H74O | 631.068 | The product derived from betulaprenyl (C45) diphosphate. This contains internal two trans isoprene residues at the w-end and the other internal isoprene residues and the a-residue are cis.(Ref. 0007) |
||||||||||||||||||||
| 22 | 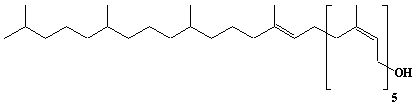 |
Glycinoprenol-9, Hexahydrononaprenol |
[2Z,6Z,10Z,14Z,18Z,22E]-3,7,11,15,19,23,27,31,35-Nonamethyl-2,6,10,14,18,22-hexatriacontahexaen-1-ol |
IIP0022 | Hiroshi Sagami |
C45H80O | 637.116 | The function is not known. |
The compound was isolated from the leaves of soybean.(Ref. 0002) |
The compound contains a phytyl residue at the w-terminal. |
||||||||||||||||||
| 23 | 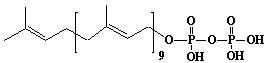 |
Decaprenyl diphosphate, trans-decaprenyl diphosphate |
[2E,6E,10E,14E,18E,22E,26E,30E,34E]-3,7,11,15,19,23,27,31,35,39-Decamethyl-2,6,10,14,18,22,26,30,34,38-tetracontadecaen-1-yl diphosphate |
IIP0023 | Hiroshi Sagami |
C50H84O7P2 | 859.145 | The product formed by the action of decaprenyl diphosphate synthase. |
||||||||||||||||||||
| 24 | 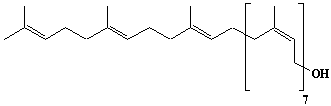 |
wt2c7-Decaprenol, cis-Decaprenol |
[2Z,6Z,10Z,14Z,18Z,22Z,26Z,30E,34E]-3,7,11,15,19,23,27,31,35,39-Decamethyl-2,6,10,14,18,22,26,30,34,38-tetracontadecaen-1-ol |
IIP0024 | Hiroshi Sagami |
C50H82O | 699.185 | The compound was isolated from bovine pituitary glands.(Ref. 0025) |
||||||||||||||||||||
| 25 | 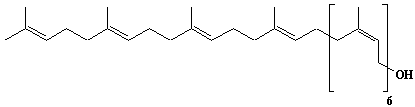 |
cis-Decaprenol, wt3c6-Decaprenol |
[2Z,6Z,10Z,14Z,18Z,22Z,26E,30E,34E]-3,7,11,15,19,23,27,31,35,39-Decamethyl-2,6,10,14,18,22,26,30,34,38-tetracontadecaen-1-ol |
IIP0025 | Hiroshi Sagami |
C50H82O | 699.185 | The compound was isolated from bovine pituitary glands.(Ref. 0025) |
||||||||||||||||||||
| 26 | 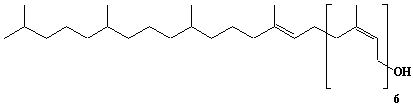 |
Glycinoprenol-10 |
[2Z,6Z,10Z,14Z,18Z,22Z,26E]-3,7,11,15,19,23,27,31,35,39-decamethyl-2,6,10,14,18,22,26-tetracontaheptaen-1-ol |
IIP0026 | Hiroshi Sagami |
C50H88O | 705.233 | The compound was isolated from the leaves of soybean.(Ref. 0002) This contains a phytyl residue at the w-terminal. |
||||||||||||||||||||
| 27 | 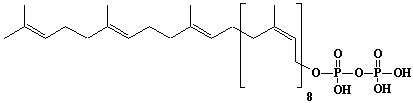 |
Undecaprenyl diphosphate, wt2c8-C55-prenyl diphosphate |
[2Z,6Z,10Z,14Z,18Z,22Z,26Z,30Z,34E,38E]-3,7,11,15,19,23,27,31,35,39,43-Undecamethyl-2,6,10,14,18,22,26,30,34,38,42-tetratetracontaundecaen-1-yl diphosphate |
IIP0027 | Hiroshi Sagami |
C55H90O7P2 | 925.246 | The compound is synthesized by the action of undecaprenyl diphosphate synthase. |
The nucleotide sequence of Micrococcus luteus undecaprenyl diphosphate synthase (Ref. 0020). |
|||||||||||||||||||
| 28 | 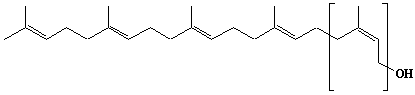 |
Undecaprenol, wt3c7-C55-prenol, Prenol-11 |
[2Z,6Z,10Z,14Z,18Z,22Z,26Z,30E,34E,38E]-3,7,11,15,19,23,27,31,35,39,43-Undecamethyl-2,6,10,14,18,22,26,30,34,38,42-tetratetracontaundecaen-1-ol |
IIP0028 | Hiroshi Sagami |
C55H90O | 767.303 | (with spectrum)(Ref. 0021) |
The effect of dolichols, polyprenols, dolichol esterified with fatty acids, and dolichyl phosphate on the structure and fluidity of model membranes was studied using 31P NMR, small-angle x-ray scattering, differential scanning calorimetry, and freeze-fracture electron microscopy. Dolichol and dolichol derivatives destabilize unsaturated phosphatidylethanolamine containing bilayer structures and promote hexagonal II phase formation. Dolichol and dolichyl-P increase the fatty acid fluidity in phosphatidylethanolamine mixtures(Ref. 0044). |
|||||||||||||||||||
| 29 | 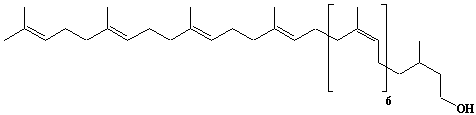 |
C55-dolichol, wt3c6-C55-dolichol, a-Dihydroundecaprenol, Dolichol-11 |
[6Z,10Z,14Z,18Z,22Z,26Z,30E,34E,38E]-3,7,11,15,19,23,27,31,35,39,43-Undecamethyl-6,10,14,18,22,26,30,34,38,42-tetratetracontadecaen-1-ol |
IIP0029 | Yoshichika Yoshioka |
Dol-11 |
C55H92O | 769.318 | The biological role of dolichol (review)(Ref. 0043). |
1H-NMR spectrum at 100MHz (with spectrum)(Ref. 0021). |
m/z=768 (with spectrum)(Ref. 0021). |
The compound was isolated from pig liver.(Ref. 0021) |
Chemical hydrogenation of plant undecaprenol(Ref. 0021). |
Dolichol-11 from pig liver was identified as cis/trans-dihydroundecaprenol in which the OH-terminal isoprene residue was saturated(Ref. 0021). The effect of dolichols, polyprenols, dolichol esterified with fatty acids, and dolichyl phosphate on the structure and fluidity of model membranes was studied using 31P NMR, small-angle x-ray scattering, differential scanning calorimetry, and freeze-fracture electron microscopy. Dolichol and dolichol derivatives destabilize unsaturated phosphatidylethanolamine containing bilayer structures and promote hexagonal II phase formation. Dolichol and dolichyl-P increase the fatty acid fluidity in phosphatidylethanolamine mixtures(Ref. 0044). |
||||||||||||||
| 30 | 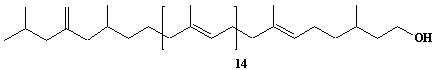 |
exo-Methylene-hexahydroprenol-18 |
exo-Methylene-hexahydroprenol-18 |
IIP0030 | Hiroshi Sagami |
C91H152O | 1262.180 | The compound was isolated from Aspergillus niger.(Ref. 0008) |
||||||||||||||||||||
| 31 | 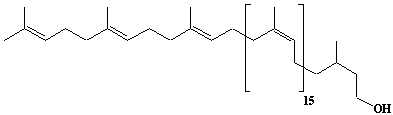 |
Dolichol-19, a-Dihydrononadecaprenol |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66E,70E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75-Nonadecamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74-hexaheptacontaoctadecaen-1-ol |
IIP0031 | Yoshichika Yoshioka |
DOL-19 |
C95H156O | 1314.255 | The compound functions as a sugar carrier lipid in the glycoprotein biosynthesis. The biological role of dolichol (review)(Ref. 0043). Content of dolichol and activity of GlcNAc-1-P (UDP-N-acetylglucosamine:dolichyl phosphate N-acetylglucosamine 1-phosphate transferase) in fibroblasts from patients with carbohydrate-deficient glycoprotein (CDG) syndrome were analyzed. The major defect of CDG syndrome is not found in the synthesys of dolichyl phosphate or GlcNAc-PP-Dol(Ref. 0061). The influence of dolichol on microsomal membrane functions: Dolichol affected microsomal membrane fluidity, but did not modify the activities of certain microsome membrane enzymes(Ref. 0062). Dolichol was covalently bound to cellular proteins with size of 5, 10, 27, 75, and >140 kDa(Ref. 0063). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dlichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
IR spectrum was observed(Ref. 0084). |
1H-NMR spectrum at 100MHz (with spectrum)(Ref. 0084/0021). The arrangement of isoprene units in pig liver dolichol was determined by 1H- and 13C-NMR spectroscopy. 1H-NMR at 200MHz, 13C-NMR at 50.1MHz (with spectra)(Ref. 0045). The head and tail sites of dolichol exibit a fast motion as exibited in the short chain isoprenols. Dolichol appears to induce a non-bilayer (isotropic) organization of phospholipid molecules in PE/PC (2:1) vesicles(Ref. 0085). |
m/z=1312 (19 isoprene units)(Ref. 0084). |
HPLC (with chromatograph)(Ref. 0046/0047/0048/0049/0050/0051). Separation of dolichol from dehydrodolichol by a simple two-plate thin-layer chromatography(Ref. 0052). Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate in human serum were made fluorometrically by HPLC. The calibration curve of dolichol standard against the internal standard showed linearity at dolichol concentrations of 10ng to 500ng(Ref. 0053). |
The compound was isolated from human kidney, ox kidney, pig heart, pig liver, rat liver, and ox intestine.(Ref. 0009). Beef and human pituitaries(Ref. 0083/0084>. Pig liver(Ref. 0021). Human tissues(Ref. 0054). Mouse testes(Ref. 0046). Rat and human tissues (Ref. 0048). Distribution in rat liver subcellular organelles (Ref. 0048). Contents in human tissues (Ref. 0055). Rat and human tissues (spleen, liver, kidney, myocardium, lung, pancreas, adrenal)(Ref. 0051). Levels of dolichol in neonatal and adult human tissues (thyroid, testis, liver, heart, kidney, lung, spleen)(Ref. 0056). Content in human serum(Ref. 0053). The effects of dietary proteins on the accumulation of dolichol in kidney, spleen, brain, and heart (Ref. 0057). Dolichol content in different regions of the rat brain (cerebral cortex, striatum, hippocampus, cerebellum, and brain stem) (Ref. 0058). Content of dolichol and retinol in isolated rat non-parenchymal liver cells(Ref. 0059). Distribution of radiolabeled dolichol in rat liver fractions (Ref. 0060). |
||||||||||||||
| 32 | 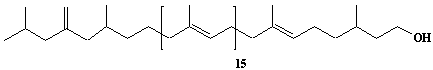 |
exo-Methylene-hexhydroprenol-19 |
exo-Methylene-hexhydroprenol-19 |
IIP0032 | Hiroshi Sagami |
C96H160O | 1330.297 | The compound was isolated from Aspergillus niger.(Ref. 0008) |
||||||||||||||||||||
| 33 | 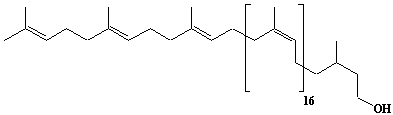 |
Dolichol-20, a-Dihydroeicosaprenol |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66Z,70E,74E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75,79-Eicosamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74,78-octacontanonadecaen-1-ol |
IIP0033 | Yoshichika Yoshioka |
Dol-20 |
C100H164O | 1382.372 | The biological role of dolichol (review)(Ref. 0043). Content of dolichol and activity of GlcNAc-1-P (UDP-N-acetylglucosamine:dolichyl phosphate N-acetylglucosamine 1-phosphate transferase) in fibroblasts from patients with carbohydrate-deficient glycoprotein (CDG) syndrome (Ref. 0061). Extraction of microsomes with n-heptane and subsequent reincorporation of dolichol were done in order to investigate the influence of dolichol on microsomal membrane functions. Dolichol affected microsomal membrane fluidity, but did not modify the activities of certain microsome membrane enzymes(Ref. 0062). Dolichol was covalently bound to cellular proteins with size of 5, 10, 27, 75, and >140 kDa(Ref. 0063). HMG-CoA reductase inhibitors induce apoptosis in mouse proximal tubular cells in primary culture(Ref. 0064). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
The arrangement of isoprene units in pig liver dolichol was determined by 1H- and 13C-NMR spectroscopy. 1H-NMR at 200MHz, 13C-NMR at 50.1MHz (with spectra)(Ref. 0045). |
HPLC (with chromatograph)(Ref. 0046/0047/0048/0049/0050/0051). Separation of dolichol from dehydrodolichol by a simple two-plate thin-layer chromatography(Ref. 0052). Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate in human serum were made fluorometrically by HPLC. The calibration curve of dolichol standard against the internal standard showed linearity at dolichol concentrations of 10ng to 500ng(Ref. 0053). |
The compound was isolated from human kidney, ox kidney, pig heart, pig liver, rat liver, and ox intestine.(Ref. 0009/0022). Human tissues (Ref. 0054). Mouse testes(Ref. 0046). Rat and human tissues (Ref. 0048). Distribution in rat liver subcellular organelles (Ref. 0048). Contents in human tissues (Ref. 0055). Rat and human tissues (spleen, liver, kidney, myocardium, lung, pancreas, adrenal)(Ref. 0051). Levels of dolichol in neonatal and adult human tissues (Ref. 0056). Content in human serum(Ref. 0053). The effects of dietary proteins on the accumulation of dolichol (Ref. 0057). Dolichol content in different regions of the rat brain (cerebral cortex, striatum, hippocampus, cerebellum, and brain stem) were analyzed from birth to the age of 2 years. The level of dolichol increased almost 100-fold during the first 10 months of life and continued to increase thereafter(Ref. 0058). Content of dolichol and retinol in isolated rat non-parenchymal liver cells(Ref. 0059). Distribution of radiolabeled dolichol in rat liver fractions (nuclei, debris, mitochondria, lysosomes, microsomes, Golgi, cytosol)(Ref. 0060). |
Incorporation of tritium from tritiated water into dolichol (Ref. 0066). Age-associated changes in dolichol metabolism (Ref. 0066). Elevated dolichol synthesis during spermatogenesis(Ref. 0046). Synthesis in liver of normal and mutant mice(Ref. 0046). Presence of abnormal amounts of dolichols (Ref. 0047). Rat liver hyperplastic nodules(Ref. 0048). Human liver cirrosis and hepatocarcinoma(Ref. 0048). Brain dolichols (Ref. 0049). Content of cerebral cortex during aging (Ref. 0049). Composition of isolated pronase-treated cytosome fractions (Ref. 0049). The content in hyperplastic nodules and developed hepatocarcinoma(Ref. 0050). Effect of age (Ref. 0067). Content in cerebral cortex of ceroid-lipofuscinosis(Ref. 0068). Age-related chnges of dolichyl phosphate content (Ref. 0051). Biosynthesis of dolichol from mevalonic acid (Ref. 0069). Farnesyl diphosphate is the common intermediate (Ref. 0070). Dolichol contents of various organs of mice(Ref. 0071). The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells (Ref. 0072). Dehydrodolichol is a common branch point intermediate(Ref. 0073). |
Metabolism of dolichol, dolichoic acid, and nordolichoic acid in MDCK and HepG2 cells (Ref. 0074). The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway (Ref. 0039). Age-dependent changes in hepatic dolichol content in rats fed either ad libitum, or subjected to restriction of daily food intake or every-other-day feeding ad libitum(Ref. 0075/0076). The effects of alcohol and glucagon on the biliary excretion of dolichols in rats (Ref. 0077). Modifications in the concentration and distribution of rat liver microsomal dolichol and dolichyl phosphate after acute ethanol adiministration (Ref. 0059). Pro-oxidant agents lead to a significant decrease of both free dolichol and dolichyl phosphate in isolated rat hepatocytes(Ref. 0059). Distribution of radiolabeled dolichol in rat liver fractions(Ref. 0060). The effect of dolichols, polyprenols, dolichol esterified with fatty acids, and dolichyl phosphate on the structure and fluidity of model membranes was studied using 31P NMR, small-angle x-ray scattering, differential scanning calorimetry, and freeze-fracture electron microscopy. (Ref. 0044). |
[0009] / [0022] / [0039] / [0043] / [0044] / [0045] / [0046] / [0047] / [0048] / [0049] / [0050] / [0051] / [0052] / [0053] / [0054] / [0055] / [0056] / [0057] / [0058] / [0059] / [0060] / [0061] / [0062] / [0063] / [0064] / [0065] / [0066] / [0067] / [0068] / [0069] / [0070] / [0071] / [0072] / [0073] / [0074] / [0075] / [0076] / [0077] |
|||||||||||||
| 34 | 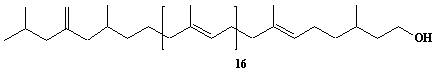 |
exo-Methylene-hexahydroprenol-20 |
exo-Methylene-hexahydroprenol-20 |
IIP0034 | Hiroshi Sagami |
C101H168O | 1398.414 | The compound was isolated from Aspergillus niger.(Ref. 0008) |
||||||||||||||||||||
| 35 | 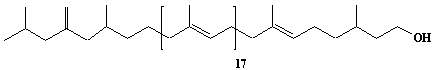 |
exo-Methylene-hexahydroprenol-21 |
exo-Methylene-hexahydroprenol-21 |
IIP0035 | Hiroshi Sagami |
C106H176O | 1466.531 | The compound was isolated from Aspergillus niger.(Ref. 0008) |
||||||||||||||||||||
| 36 | 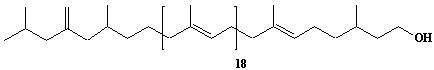 |
exo-Methylene-hexhydroprenol-22 |
exo-Methylene-hexhydroprenol-22 |
IIP0036 | Hiroshi Sagami |
C111H184O | 1534.648 | The compound was isolated from Aspergillus niger.(Ref. 0008) |
||||||||||||||||||||
| 37 | 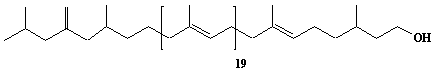 |
exo-Methylene-hexhydroprenol-23 |
exo-Methylene-hexhydroprenol-23 |
IIP0037 | Hiroshi Sagami |
C116H192O | 1602.765 | The compound was isolated from Aspergillus niger.(Ref. 0008) |
||||||||||||||||||||
| 38 | 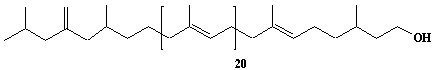 |
exo-Methylene-hexahydroprenol-24 |
exo-Methylene-hexahydroprenol-24 |
IIP0038 | Hiroshi Sagami |
C121H200O | 1670.882 | The compound was isolated from Aspergillus niger.(Ref. 0008) |
||||||||||||||||||||
| 39 | 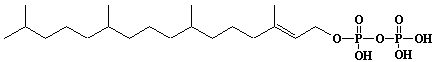 |
Phytyl diphosphate |
[2E]-3,7,11,15-Tetramethyl-2-hexadecen-1-yl diphosphate |
IIP0039 | Hiroshi Sagami |
C20H42O7P2 | 456.491 | The compound is formed from geranylgeranyl diphosphate by the action of geranylgeranyl reductase. |
The nucleotide sequence of geranylgeranyl reductase is shown in (Ref. 0027). |
|||||||||||||||||||
| 40 | 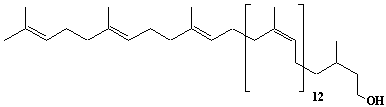 |
Dolichol-16, a-Dihydrohexadecaprenol |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54E,58E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63-Hexadecamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62-tetrahexacontapentadecaen-1-ol |
IIP0040 | Yoshichika Yoshioka |
Dol-16 |
C80H132O | 1109.903 | The biological role of dolichol (review)(Ref. 0043). |
Separation of dolichol from dehydrodolichol by a simple two-plate thin-layer chromatography(Ref. 0052). |
The compound was found as prenol in Schizosaccharomyces pombe(Ref. 0028). Dolichol content in different regions of the rat brain (cerebral cortex, striatum, hippocampus, cerebellum, and brain stem) were analyzed from birth to the age of 2 years. The level of dolichol increased almost 100-fold during the first 10 months of life and continued to increase thereafter(Ref. 0058). |
Incorporation of tritium from tritiated water into dolichol in the kidneys and liver of mice as a function of age(Ref. 0066). Age-associated changes in dolichol metabolism in the kidneys and liver of mice(Ref. 0066). Dolichol contents of various organs of mice used as a model for Niemann-Pick's type C disease (liver, spleen, kidney, brain, testis, lung, adipose tissue, heart, intestine, skeletal muscle). Contents in subfractions isolated from the liver of mice used as a model for NPC (mitocondria, lysosomes, microsomes)(Ref. 0071). The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capasity were as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and dolichol were not inhibitors(Ref. 0039). |
||||||||||||||||
| 41 | 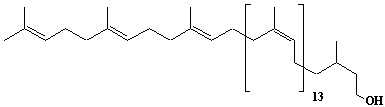 |
Dolichol-17, a-Dihydroheptadecaprenol |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58E,62E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67-Heptadecamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66-octahexacontahexadecaen-1-ol |
IIP0041 | Yoshichika Yoshioka |
Dol-17 |
C85H140O | 1178.020 | The biological role of dolichol (review) (Ref. 0043). Content of dolichol and activity of GlcNAc-1-P (UDP-N-acetylglucosamine:dolichyl phosphate N-acetylglucosamine 1-phosphate transferase) in fibroblasts from patients with carbohydrate-deficient glycoprotein (CDG) syndrome (Ref. 0061). Extraction of microsomes with n-heptane and subsequent reincorporation of dolichol were done in order to investigate the influence of dolichol on microsomal membrane functions. Dolichol affected microsomal membrane fluidity, but did not modify the activities of certain microsome membrane enzymes(Ref. 0062). Dolichol was covalently bound to cellular proteins with size of 5, 10, 27, 75, and >140 kDa(Ref. 0063). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dlichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
HPLC (with chromatograph)(Ref. 0046/0048/0050/0051). Separation of dolichol from dehydrodolichol by a simple two-plate thin-layer chromatography(Ref. 0052). Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate in human serum were made fluorometrically by HPLC. The calibration curve of dolichol standard against the internal standard showed linearity at dolichol concentrations of 10ng to 500ng(Ref. 0053). |
The compound was found as prenol in Schizosaccharomyces pombe(Ref. 0028). Human tissues (Ref. 0054). Mouse testes(Ref. 0046). Rat and human tissues(Ref. 0048). In rat liver, outer mitochondrial and Golgi membranes, lysosomes and plasmamembranes contain considerable amounts of dolichol(Ref. 0048). Contents in human tissues (Ref. 0055). Rat and human tissues(Ref. 0051). Levels of dolichol in neonatal and adult human tissues (Ref. 0056). Content in human serum(Ref. 0053). The effects of dietary proteins on the accumulation of dolichol in kidney, spleen, brain, and heart were studied. There were increases in dolichol contents in rat tissues with age. The contents of kidney and spleen were influenced by the diet(Ref. 0057). Dolichol content in different regions of the rat brain were analyzed from birth to the age of 2 years. The level of dolichol increased almost 100-fold during the first 10 months of life and continued to increase thereafter(Ref. 0058). Content of dolichol and retinol in isolated rat non-parenchymal liver cells(Ref. 0059). Distribution of radiolabeled dolichol in rat liver fractions (Ref. 0060). |
Incorporation of tritium from tritiated water into dolichol (Ref. 0066). Age-associated changes in dolichol metabolism (Ref. 0066). Elevated dolichol synthesis during spermatogenesis(Ref. 0046). Synthesis in liver of normal and mutant mice(Ref. 0046). Rat liver hyperplastic nodules(Ref. 0048). Human liver cirrosis and hepatocarcinoma(Ref. 0048). The content in hyperplastic nodules of rat liver (Ref. 0050). Effect of age on the content in brain, liver, spleen, kidney, and testis of the rat(Ref. 0067). Content in cerebral cortex of ceroid-lipofuscinosis(Ref. 0068). Age-related chnges of dolichyl phosphate content (Ref. 0051). Farnesyl diphosphate is the common intermediate (Ref. 0070). Dolichol contents of various organs of mice (Ref. 0071). The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells (Ref. 0072). Dehydrodolichol is a common branch point intermediate (Ref. 0073). Metabolism of dolichol, dolichoic acid, and nordolichoic acid (Ref. 0074). The potential for feedback inhibition by isoprene intermediates (Ref. 0039). |
Age-dependent changes in hepatic dolichol content in rats fed either ad libitum, or subjected to restriction of daily food intake or every-other-day feeding ad libitum (Ref. 0075/0076). The effects of alchol and glucagon on the biliary excretion of dolichols in rats were investigated. Chronic ethanol treatment decreased both biliary dolichol and beta-hexosaminidase excretion. (Ref. 0077). Modifications in the concentration and distribution of rat liver microsomal dolichol and dolichyl phosphate after acute ethanol adiministration (6 g/kg body weight) were investigated. Between 3 and 24 hr after ethanol administration, the microsomal dolichyl phosphate concentration was significantly lower than in control rats. An earlier and more marked reduction of total dolichol was observed in the Golgi apparatus(Ref. 0059). Pro-oxidant agents lead to a significant decrease of both free dolichol and dolichyl phosphate in isolated rat hepatocytes(Ref. 0059). Distribution of radiolabeled dolichol in rat liver fractions(Ref. 0060). |
|||||||||||||||
| 42 |  |
Squalene |
2,6,10,15,19,23-Hexamethyl-2,6,10,14,18,22-tetraacosahexaene |
IIP0042 | Hiroshi Sagami |
C30H50 | 410.718 | The compound was isolated from Halobacterium cutirubrum with its di- and tetrahydrosqualenes(Ref. 0029). |
Molecular cloning of the yeast gene for squalene synthetase(Ref. 0030) |
|||||||||||||||||||
| 43 | 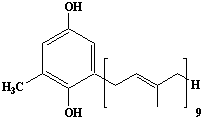 |
2-Demethylplastoquinol, 2-Methyl-6-nonaprenylbenzoquinol |
IIP0043 | Hiroshi Sagami |
C52H80O2 | 737.190 | The compound is formed from homogentisate and nonaprenyl pyrophosphate by the action of homogentisate decarboxylase-nonaprenyltransferase(Ref. 0031). |
|||||||||||||||||||||
| 44 | 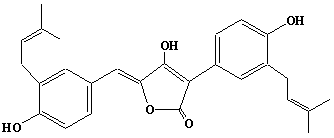 |
Aspulvinone H |
IIP0044 | Hiroshi Sagami |
C27H28O5 | 432.508 | The compound is formed from Aspulvinone E and dimethylallyl diphosphate by the action of dimethylallyl pyrophosphate:aspulvinone dimethylallyltransferase.(Ref. 0032) |
|||||||||||||||||||||
| 45 | 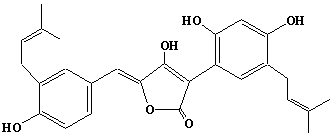 |
Aspulvinone J |
IIP0045 | Hiroshi Sagami |
C27H28O3 | 400.509 | The compound is formed from Aspulvinone G and dimethylallyl diphosphate by the action of dimethylallyl pyrophosphate:aspulvinone dimethylallyltransferase.(Ref. 0032) |
|||||||||||||||||||||
| 46 | 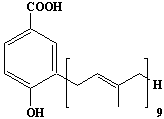 |
3-Nonaprenyl-4-hydroxybenzoate |
IIP0046 | Hiroshi Sagami |
C52H78O3 | 751.174 | The compound is formed from 4-hydroxybenzoate derived from tyrosine metabolism and nonaprenyl pyrophosphate derived from mevalonic acid by the action of 4-hydroxybenzoate polyprenyltransferase (Ref. 0033).This is the major product of the enzymatic reaction in rat liver. |
|||||||||||||||||||||
| 47 | 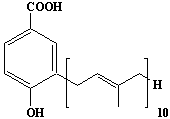 |
3-Decaprenyl-4-hydroxybenzoate |
IIP0047 | Hiroshi Sagami |
C57H86O3 | 819.291 | The compound is formed from 4-hydroxybenzoate derived from tyrosine metabolism and decaprenyl pyrophosphate derived from mevalonic acid by the action of 4-hydroxybenzoate polyprenyltransferase (Ref. 0033). This is the major product of the enzymatic reaction in human and guinea pig. |
|||||||||||||||||||||
| 48 | 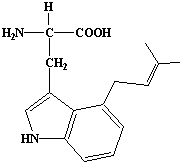 |
4-Dimethylallyltryptophan |
IIP0048 | Hiroshi Sagami |
C16H20O2N2 | 272.342 | The compound is formed from tryptophan and dimethylallyl pyrophosphate by the action of dimethylallylpyrophosphate:L-tryptophan dimethylallyltransferase(Ref. 0034). |
|||||||||||||||||||||
| 49 | 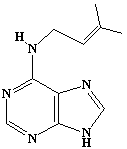 |
Isopentenyladenine |
IIP0049 | Hiroshi Sagami |
C10H13N5 | 203.244 | The compound acts as a mediator of mevalonate-regulated DNA replication (Ref. 0035). |
Isolation procedures are described in (Ref. 0036). |
||||||||||||||||||||
| 50 | 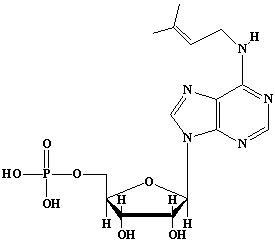 |
Dimethylallyladenosine5'-phosphate, N6-(d2-isopentenyl)adenosine5'-phosphate, N6-(g,g-dimethylallyl)adenosine5'-phosphate |
IIP0050 | Hiroshi Sagami |
iPA-5'P, iPeAde-5'P |
C15H22O7N5P1 | 415.338 | The compound is formed from 5'-AMP and dimethylallyl pyrophosphate by the action of dimethylallyladenosine 5'-AMP dimethylallyltransferase(Ref. 0037). |
||||||||||||||||||||
| 51 |  |
Farnesol |
3,7,11-Trimethyl-2,6,10-dodecatrien-1-ol |
IIP0051 | Yoshichika Yoshioka |
FOH |
C15H26O | 222.366 | Farnesol inhibited growth of leukemia cells (CEM-C1) without causing cell lysis.(Ref. 0086) Farnesol inhibited phosphatidylcholine biosynthesis in cultured cells dy decreasing cholinephosphotransferase activity.(Ref. 0087) In HeLa cells farnesol caused translocation of PKC from membrane fraction to cytosol after 1hr of incubation and also prevented PMA-stimulated induction of PKC translocation from cytosol to membranes.(Ref. 0088) |
120 degree Celsius (at 0.2 mmHg) |
The 2H-labeled farnesol gave rise to 2H NMR powder patterns interpretable in terms of quadrupole splittings and splin-lattice relaxation times. Spin-lattice relaxation time measurements revealed high rates of motion for geraniol relative to cholesterol in similar membrane hosts and revealed correlation times close to the fatty acyl methyl termini in phosphatidylcholine.(Ref. 0038) |
Pig liver(Ref. 0021). |
Incorporation of mevalonic acid-derived materials into proteins was studied with extremely halophilic archaebacteria, Halobacterium halobium and Halobacterium cutirubrum. The amount of farnesylated proteins was very small in H. halobium(Ref. 0089). The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capasity were as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and dolichol were not inhibitors(Ref. 0039). |
Tree dimansional structure(Ref. 0090). |
||||||||||||||
| 52 |  |
Geranylfarnesol |
3,7,11,15,18-Pentamethyl-2,6,10,14,18-eicosapentaen-1-ol |
IIP0052 | Yoshichika Yoshioka |
GFOH |
C25H42O | 358.600 | Geranylfarnesol had a potent apoptosis-inducing activity in HL-60 cells.(Ref. 0040) |
|||||||||||||||||||
| 53 | 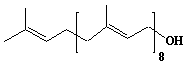 |
Solanesol, Prenol-9, Nonaprenol |
[2E,6E,10E,14E,18E,22E,26E,30E]-3,7,11,15,19,23,27,31,35-Nonamethyl-2,6,10,14,18,22,26,30,34-hexatriacontanonaene-1-ol |
IIP0053 | Yoshichika Yoshioka |
C45H74O | 631.068 | The 2H-labeled solanesol gave rise to 2H NMR powder patterns interpretable in terms of quadrupole splittings and splin-lattice relaxation times. Spin-lattice relaxation time measurements revealed high rates of motion for geraniol relative to cholesterol in similar membrane hosts and revealed correlation times close to the fatty acyl methyl termini in phosphatidylcholine.(Ref. 0038) |
||||||||||||||||||||
| 54 | 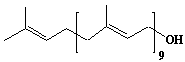 |
Decaprenol, Prenol-10 |
[2E,6E,10E,14E,18E,22E,26E,30E,34E]-3,7,11,15,19,23,27,31,35,39-Decamethyl-2,6,10,14,18,22,26,30,34,38-tetracontadecaene-1-ol |
IIP0054 | Yoshichika Yoshioka |
C50H82O | 699.185 | |||||||||||||||||||||
| 55 | 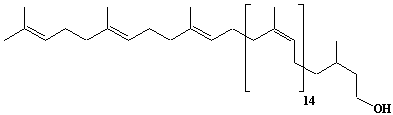 |
Dolichol-18, a-Dihydrooctadecaprenol |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62E,66E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71-Octadecamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70-doheptacontaheptadecaen-1-ol |
IIP0055 | Yoshichika Yoshioka |
Dol-18 |
C90H148O | 1246.138 | The biological role of dolichol (review)(Ref. 0043). Content of dolichol and activity of GlcNAc-1-P (UDP-N-acetylglucosamine:dolichyl phosphate N-acetylglucosamine 1-phosphate transferase) in fibroblasts from patients with carbohydrate- deficient- glycoprotein (CDG) syndrome were analyzed. The major defect of CDG syndrome is not found in the synthesys of dolichyl phosphate or GlcNAc-PP-Dol(Ref. 0061). Extraction of microsomes with n-heptane and subsequent reincorporation of dolichol were done in order to investigate the influence of dolichol on microsomal membrane functions. Dolichol affected microsomal membrane fluidity, but did not modify the activities of certain microsome membrane enzymes(Ref. 0062). Dolichol was covalently bound to cellular proteins with size of 5, 10, 27, 75, and >140 kDa(Ref. 0063). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dlichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
The arrangement of isoprene units in pig liver dolichol was determined by 1H- and 13C-NMR spectroscopy. 1H-NMR at 200MHz, 13C-NMR at 50.1MHz (with spectra)(Ref. 0045). |
HPLC (with chromatograph)(Ref. 0046/0047/0048/0049/0050/0051). Separation of dolichol from dehydrodolichol by a simple two-plate thin-layer chromatography(Ref. 0052). Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate in human serum were made fluorometrically by HPLC. The calibration curve of dolichol standard against the internal standard showed linearity at dolichol concentrations of 10ng to 500ng(Ref. 0053). |
Human tissues (testes, adrenal ,liver, thyroid, pancreas, prostate, heart, kidney, spleen, lung)(Ref. 0054). Mouse testes(Ref. 0046). Rat and human tissues (heart, kidney, liver, lung, intestine, testis, brain, spleen, pituitary)(Ref. 0048). Distribution in rat liver subcellular organelles (outer mitochondrial and Golgi membranes, lysosomes and plasmamembranes contain considerable amount)(Ref. 0048). Contents in human tissues (adrenal, aorta, brain, colon, fatty tissue, heart, kidney, liver, lung, muscle, ovary, pancreas, pituitary gland, placenta, prostate, small intestine, spleen, testis, tyroid grand, uterus, and stomach)(Ref. 0055). Rat and human tissues (spleen, liver, kidney, myocardium, lung, pancreas, adrenal)(Ref. 0051). Levels of dolichol in neonatal and adult human tissues (thyroid, testis, liver, heart, kidney, lung, spleen)(Ref. 0056). Content in human serum(Ref. 0053). The effects of dietary proteins on the accumulation of dolichol in kidney, spleen, brain, and heart were studied. There were increases in dolichol contents in rat tissues with age. The contents of kidney and spleen were influenced by the diet(Ref. 0057). Dolichol content in different regions of the rat brain (cerebral cortex, striatum, hippocampus, cerebellum, and brain stem) were analyzed from birth to the age of 2 years. The level of dolichol increased almost 100-fold during the first 10 months of life and continued to increase thereafter(Ref. 0058). Content of dolichol and retinol in isolated rat non-parenchymal liver cells(Ref. 0106). Distribution of radiolabeled dolichol in rat liver fractions (nuclei, debris, mitochondria, lysosomes, microsomes, Golgi, cytosol)(Ref. 0060). |
Incorporation of tritium from tritiated water into dolichol in the kidneys and liver of mice as a function of age(Ref. 0066). Age-associated changes in dolichol metabolism in the kidneys and liver of mice(Ref. 0066). Elevated dolichol synthesis in mouse testes during spermatogenesis(Ref. 0046). Synthesis in liver of normal and mutant mice(Ref. 0046). Presence of abnormal amounts of dolichols in the urinary sediment of Batten disease (neuronal ceroid lipofuscinosis) patients(Ref. 0047). Rat liver hyperplastic nodules(Ref. 0048). Human liver cirrosis and hepatocarcinoma(Ref. 0048). Brain dolichols in neuronal ceroid lipofuscinosis, epilepsy, Tay-Sachs desease, Juvenile Huntington disease, Alzheimer's disease, Pick disease, Astrocytoma(grade III-IV)(Ref. 0049). Content of cerebral cortex during aging (human & rat)(Ref. 0049). Composition of isolated pronase-treated cytosome fractions from neuronal ceroid-lipofuscinosis patients(Ref. 0049). The dolichol content of hyperplastic nodules in rat liver was four times that of normal. In developed hepatocarcinoma, the amount of dolichol was doubled(Ref. 0050). Effect of age on the content in brain, liver, spleen, kidney, and testis of the rat(Ref. 0067). Absorption and distribution of dolichol intubated into rats(Ref. 0091). Content in cerebral cortex of ceroid-lipofuscinosis(Ref. 0068). Age-related chnges of dolichyl phosphate content in rat and human tissues (Ref. 0051). Biosynthesis of dolichol from mevalonic acid was measured in non-malignant and malignant cultured human lymphocytes, freshly isolated human mononuclear leucocytes and in cultured human hepatoma cells(Ref. 0069). Formation of dolichol from dehydrodolichol is catalyzed by NADPH-dependent reductase localized in microsomes of rat liver. Dolichol was syntesized from isopentenyl diphosphate and Z,E,E-geranylgeranyl diphosphate. Neither dolichyl diphosphate nor dolichyl phosphate was detected(Ref. 0092). Farnesyl diphosphate is the common intermediate at the branch point for the synthesis of geranylgeranylated proteins as well as cholesterol, ubiquinone, dolichol, and farnesylated proteins(Ref. 0070). Dolichol contents of various organs of mice used as a model for Niemann-Pick's type C disease . Contents in subfractions isolated from the liver of mice used as a model for NPC (mitocondria, lysosomes, microsomes)(Ref. 0071). |
The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of dolichol and acitivated the oligosaccharyltransferase(Ref. 0072). Dehydrodolichol is a common branch point intermediate in the biosynthetic pathways leading to dolichal and dolichol(Ref. 0073). Metabolism of dolichol, dolichoic acid, and nordolichoic acid in MDCK and HepG2 cells were investigated. Dolichoic acid was broken down by alpha-oxidation and also converted to a mixture of polar compounds. No oxidative breakdown of dolichol was found(Ref. 0074). The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capasity were as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and dolichol were not inhibitors(Ref. 0039). Age-dependent changes in hepatic dolichol content in rats fed either ad libitum, or subjected to restriction of daily food intake or every-other-day feeding ad libitum (antiaging dietary restriction). Both types of antiaging dietary restriction fully prevented the age-associated accumulation of dolichol in the liver and had a much smaller preventive effect in the kidney(Ref. 0075/0076). The effects of alcohol and glucagon on the biliary excretion of dolichols in rats were investigated. Chronic ethanol treatment decreased both biliary dolichol and beta-hexosaminidase excretion. The absolute amount of dolichol excreted into the bile correlated highly significantly with the absolute amount of biliary beta-hexosaminidase. Biliary dolichols are derived from hepatic lysosomes(Ref. 0077). Modifications in the concentration and distribution of rat liver microsomal dolichol and dolichyl phosphate after acute ethanol adiministration (6 g/kg body weight) were investigated. Between 3 and 24 hr after ethanol administration, the microsomal dolichyl phosphate concentration was significantly lower than in control rats. An earlier and more marked reduction of total dolichol was observed in the Golgi apparatus(Ref. 0059). Pro-oxidant agents lead to a significant decrease of both free dolichol and dolichyl phosphate in isolated rat hepatocytes(Ref. 0059). Distribution of radiolabeled dolichol in rat liver fractions(Ref. 0060). |
[0039] / [0043] / [0045] / [0046] / [0047] / [0048] / [0049] / [0050] / [0051] / [0052] / [0053] / [0054] / [0055] / [0056] / [0057] / [0058] / [0059] / [0060] / [0061] / [0062] / [0063] / [0065] / [0066] / [0067] / [0068] / [0069] / [0070] / [0071] / [0072] / [0073] / [0074] / [0075] / [0076] / [0077] / [0091] / [0092] / [0106] |
|||||||||||||
| 56 | 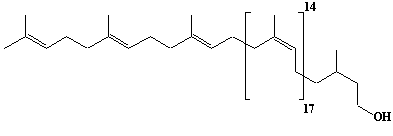 |
Dolichol-21, a-Dihydroheneicosaprenol |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66Z,70Z,74E,78E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75,79,83-Henicosamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74,78,82-tetraoctacontaeicosaen-1-ol |
IIP0056 | Yoshichika Yoshioka |
Dol-21 |
C105H172O | 1450.489 | The biological role of dolichol (review)(Ref. 0043). Content of dolichol and activity of GlcNAc-1-P (UDP-N-acetylglucosamine:dolichyl phosphate N-acetylglucosamine 1-phosphate transferase) in fibroblasts from patients with carbohydrate-deficient-glycoprotein (CDG) syndrome were analyzed. The major defect of CDG syndrome is not found in the synthesys of dolichyl phosphate or GlcNAc-PP-Dol(Ref. 0061). Extraction of microsomes with n-heptane and subsequent reincorporation of dolichol were done in order to investigate the influence of dolichol on microsomal membrane functions. Dolichol affected microsomal membrane fluidity, but did not modify the activities of certain microsome membrane enzymes(Ref. 0062). Dolichol was covalently bound to cellular proteins with size of 5, 10, 27, 75, and >140 kDa(Ref. 0063). HMG-CoA reductase inhibitors induce apoptosis in mouse proximal tubular cells in primary culture. Geranylgeranylpyrophosphate and mevalonate completely reversed the effect of the inhibitors, while farnesylpyrophosphate partially reversed it and dolichol had no effect(Ref. 0064). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dlichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
HPLC (with chromatograph)(Ref. 0046/0047/0048/0049/0050/0051). Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate in human serum were made fluorometrically by HPLC. The calibration curve of dolichol standard against the internal standard showed linearity at dolichol concentrations of 10ng to 500ng(Ref. 0053). |
Human tissues (testes, adrenal, liver, thyroid, pancreas, prostate, heart, kidney, spleen, lung)(Ref. 0054). Mouse testes(Ref. 0046). Rat and human tissues (heart, kidney, liver, lung, intestine, testis, brain, spleen, pituitary)(Ref. 0048). Distribution in rat liver subcellular organelles (outer mitochondrial and Golgi membranes, lysosomes and plasmamembranes contain considerable amount)(Ref. 0048). Contents in human tissues (adrenal, aorta, brain, colon, fatty tissue, heart, kidney, liver, lung, muscle, ovary, pancreas, pituitary gland, placenta, prostate, small intestine, spleen, testis, tyroid grand, uterus, and stomach)(Ref. 0055). Rat and human tissues (spleen, liver, kidney, myocardium, lung, pancreas, adrenal)(Ref. 0051). Levels of dolichol in neonatal and adult human tissues (thyroid, testis, liver, heart, kidney, lung, spleen)(Ref. 0056). Content in human serum(Ref. 0053). The effects of dietary proteins on the accumulation of dolichol in kidney, spleen, brain, and heart were studied. There were increases in dolichol contents in rat tissues with age. The contents of kidney and spleen were influenced by the diet(Ref. 0057). Dolichol content in different regions of the rat brain (cerebral cortex, striatum, hippocampus, cerebellum, and brain stem) were analyzed from birth to the age of 2 years. The level of dolichol increased almost 100-fold during the first 10 months of life and continued to increase thereafter(Ref. 0058). Content of dolichol and retinol in isolated rat non-parenchymal liver cells(Ref. 0106). |
Incorporation of tritium from tritiated water into dolichol in the kidneys and liver of mice as a function of age(Ref. 0066). Age-associated changes in dolichol metabolism in the kidneys and liver of mice(Ref. 0066). Elevated dolichol synthesis in mouse testes during spermatogenesis(Ref. 0046). Synthesis in livers of normal and mutant mice(Ref. 0046). Presence of abnormal amounts of dolichols in the urinary sediment of Batten disease (neuronal ceroid lipofuscinosis) patients(Ref. 0047). Rat liver hyperplastic nodules(Ref. 0048). Human liver cirrosis and hepatocarcinoma(Ref. 0048). Brain dolichols in neuronal ceroid lipofuscinosis, epilepsy, Tay-Sachs desease, Juvenile Huntington disease, Alzheimer's disease, Pick disease, Astrocytoma(grade III-IV)(Ref. 0049). Content of cerebral cortex during aging (human & rat)(Ref. 0049). Composition of isolated pronase-treated cytosome fractions from neuronal ceroid-lipofuscinosis patients(Ref. 0049). The content in hyperplastic nodules of rat liver increased four times in the homoginate and six times in the microsomes. In developed hepatocarcinoma, the amount was doubled(Ref. 0050). Effect of age on the content in brain, liver, spleen, kidney, and testis of the rat(Ref. 0067). Content in cerebral cortex of ceroid-lipofuscinosis(Ref. 0068). Age-related chnges of dolichyl phosphate content in rat and human tissues (Ref. 0051). Biosynthesis of dolichol from mevalonic acid was measured in non-malignant and malignant cultured human lymphocytes, freshly isolated human mononuclear leucocytes and in cultured human hepatoma cells(Ref. 0069). Dolichol contents of various organs of mice used as a model for Niemann-Pick's type C disease . Contents in subfractions isolated from the liver of mice used as a model for NPC (Ref. 0071). The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of dolichol and acitivated the oligosaccharyltransferase(Ref. 0072). Age-dependent changes in hepatic dolichol content in rats fed either ad libitum, or subjected to restriction of daily food intake or every-other-day feeding ad libitum (antiaging dietary restriction). Both types of antiaging dietary restriction fully prevented the age-associated accumulation of dolichol in the liver and had a much smaller preventive effect in the kidney(Ref. 0075/0076). |
The effects of alchol and glucagon on the biliary excretion of dolichols in rats were investigated. Chronic ethanol treatment decreased both biliary dolichol and beta-hexosaminidase excretion. The absolute amount of dolichol excreted into the bile correlated highly significantly with the absolute amount of biliary beta-hexosaminidase. Biliary dolichols are derived from hepatic lysosomes(Ref. 0077). Modifications in the concentration and distribution of rat liver microsomal dolichol and dolichyl phosphate after acute ethanol adiministration (6 g/kg body weight) were investigated. Between 3 and 24 hr after ethanol administration, the microsomal dolichyl phosphate concentration was significantly lower than in control rats. An earlier and more marked reduction of total dolichol was observed in the Golgi apparatus(Ref. 0059). Pro-oxidant agents lead to a significant decrease of both free dolichol and dolichyl phosphate in isolated rat hepatocytes(Ref. 0059). |
|||||||||||||||
| 57 | 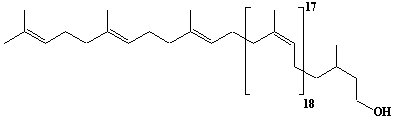 |
Dolichol-22, a-Dihydrodocosaprenol |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66Z,70Z,74Z,78E,82E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75,79,83,87-Docosamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74,78,82,86-octaoctacontahenicosaen-1-ol |
IIP0057 | Yoshichika Yoshioka |
Dol-22 |
C110H180O | 1518.606 | The biological role of dolichol (review)(Ref. 0043). Extraction of microsomes with n-heptane and subsequent reincorporation of dolichol were done in order to investigate the influence of dolichol on microsomal membrane functions. Dolichol affected microsomal membrane fluidity, but did not modify the activities of certain microsome membrane enzymes(Ref. 0062). HMG-CoA reductase inhibitors induce apoptosis in mouse proximal tubular cells in primary culture. Geranylgeranylpyrophosphate and mevalonate completely reversed the effect of the inhibitors, while farnesylpyrophosphate partially reversed it and dolichol had no effect(Ref. 0064). |
Human tissues (testes, adrenal, liver, thyroid, pancreas, prostate, heart, kidney, spleen, lung)(Ref. 0054). Human tissues (liver, testis, spleen, pituitary)(Ref. 0048). Rat and human tissues (spleen, liver, kidney, myocardium, lung, pancreas, adrenal)(Ref. 0051). The effects of dietary proteins on the accumulation of dolichol in kidney, spleen, brain, and heart were studied. There were increases in dolichol contents in rat tissues with age. The contents of kidney and spleen were influenced by the diet(Ref. 0057). Dolichol content in different regions of the rat brain (cerebral cortex, striatum, hippocampus, cerebellum, and brain stem) were analyzed from birth to the age of 2 years. The level of dolichol increased almost 100-fold during the first 10 months of life and continued to increase thereafter(Ref. 0058). |
Rat liver hyperplastic nodules(Ref. 0048). Human liver cirrosis and hepatocarcinoma (Ref. 0048). Age-related chnges of dolichyl phosphate content in rat and human tissues (spleen, liver, kidney, heart, lung, pancreas, adrenal)(Ref. 0051). Dolichol contents of various organs of mice used as a model for Niemann-Pick's type C disease (liver, spleen, kidney, brain, testis, lung, adipose tissue, heart, intestine, skeletal muscle). Contents in subfractions isolated from the liver of mice used as a model for NPC (mitocondria, lysosomes, microsomes)(Ref. 0071). |
|||||||||||||||||
| 58 | 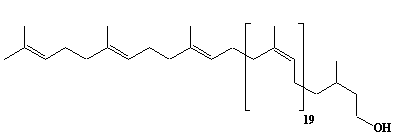 |
Dolichol-23, a-Dihydrotricosaprenol |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66Z,70Z,74Z,78Z,82E,86E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75,79,83,87,91-Tricosamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74,78,82,86,90-dononacontadocosaen-1-ol |
IIP0058 | Yoshichika Yoshioka |
Dol-23 |
C115H188O | 1586.723 | The biological role of dolichol (review)(Ref. 0043). HMG-CoA reductase inhibitors induce apoptosis in mouse proximal tubular cells in primary culture. Geranylgeranylpyrophosphate and mevalonate completely reversed the effect of the inhibitors, while farnesylpyrophosphate partially reversed it and dolichol had no effect(Ref. 0064). |
1H-NMR spectrum at 100MHz (with spectrum)(Ref. 0048). |
Bovine pituitary glands(Ref. 0048). |
|||||||||||||||||
| 59 | 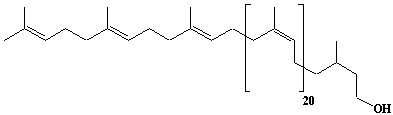 |
Dolichol-24, a-Dihydrotetracosaprenol |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66Z,70Z,74Z,78Z,82Z,86E,90E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75,79,83,87,91,95-Tetracosamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74,78,82,86,90,94-hexanonacontatricosaen-1-ol |
IIP0059 | Yoshichika Yoshioka |
Dol-24 |
C120H196O | 1654.840 | HMG-CoA reductase inhibitors induce apoptosis in mouse proximal tubular cells in primary culture. Geranylgeranylpyrophosphate and mevalonate completely reversed the effect of the inhibitors, while farnesylpyrophosphate partially reversed it and dolichol had no effect(Ref. 0064). |
The effect of dolichols, polyprenols, dolichol esterified with fatty acids, and dolichyl phosphate on the structure and fluidity of model membranes was studied using 31P NMR, small-angle x-ray scattering, differential scanning calorimetry, and freeze-fracture electron microscopy. Dolichol and dolichol derivatives destabilize unsaturated phosphatidylethanolamine containing bilayer structures and promote hexagonal II phase formation. Dolichol and dolichyl-P increase the fatty acid fluidity in phosphatidylethanolamine mixtures(Ref. 0044). |
||||||||||||||||||
| 60 | 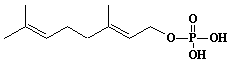 |
Geranyl monophosphate |
[2E]-3,7-Dimethyl-2,6-octadien-1-yl monophosphate |
IIP0060 | Yoshichika Yoshioka |
GOP |
C10H19O4P | 234.229 | Metabolic labeling of rat liver slices with mevalonic acid revealed the accumulation of E,E,E-geranylgeranyl (di)phosphate as well as dolichyl (di)phosphate (C85 and C90) and dehydrodolichol (C85 and C90), but no accumulation of Z,E,E-geranylgeranyl (di)phosphate or E,E-farnesyl (di)phosphate was detected(Ref. 0026). |
|||||||||||||||||||
| 61 |  |
Farnesyl monophosphate |
[2E,6E]-3,7,11-Trimethyl-2,6,10-dodecatrien-1-yl monophosphate |
IIP0061 | Yoshichika Yoshioka |
FOP |
C15H27O4P | 302.346 | Metabolic labeling of rat liver slices with mevalonic acid revealed the accumulation of E,E,E-geranylgeranyl (di)phosphate as well as dolichyl (di)phosphate (C85 and C90) and dehydrodolichol (C85 and C90), but no accumulation of Z,E,E-geranylgeranyl (di)phosphate or E,E-farnesyl (di)phosphate was detected(Ref. 0026). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dlichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
|||||||||||||||||||
| 62 | 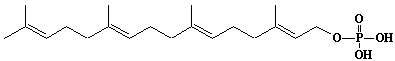 |
Geranylgeranyl monophosphate |
[2E,6E,10E]-3,7,11,15-Tetramethyl-2,6,10,14-hexadecatetraen-1-yl monophosphate |
IIP0062 | Yoshichika Yoshioka |
GGOP |
C20H35O4P | 370.463 | Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dlichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
|||||||||||||||||||
| 63 | 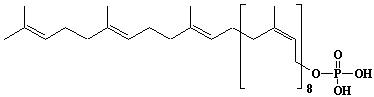 |
Prenyl-11 monophosphate, Undecaprenyl monophosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30E,34E,38E]-3,7,11,15,19,23,27,31,35,39,43-Undecamethyl-2,6,10,14,18,22,26,30,34,38,42-tetratetracontaundecaen-1-yl monophosphate |
IIP0063 | Yoshichika Yoshioka |
C55H91O4P | 847.282 | The head and tail sites of undecaprenol exibit a fast motion as exibited in the short chain isoprenols. Undecaprenol appears to induce a non-bilayer (isotropic) organization of phospholipid molecules in PE/PC (2:1) vesicles(Ref. 0085). |
||||||||||||||||||||
| 64 | 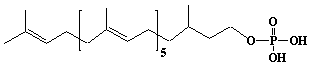 |
Dihydroprenyl-7 monophosphate, a-Dihydroheptaprenyl monophosphate |
[6E,10E,14E,18E,22E]-3,7,11,15,19,23,27-Heptamethyl-6,10,14,18,22,26-octacosahexaen-1-yl monophosphate |
IIP0064 | Yoshichika Yoshioka |
C35H61O4P | 576.830 | Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dlichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
||||||||||||||||||||
| 65 | 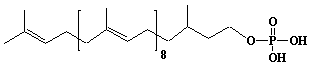 |
Dihydroprenyl-10 monophosphate, a-Dihydrodecaprenyl monophosphate |
[6E,10E,14E,18E,22E,26E,30E,34E]-3,7,11,15,19,23,27,31,35,39-Decamethyl-6,10,14,18,22,26,30,34,38-Tetracontanonaen-1-yl monophosphate |
IIP0065 | Yoshichika Yoshioka |
C50H85O4P | 781.181 | Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dolichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
The dynamics of dolichyl phosphate molecules in membranes can be regulated by transmembrane electrical potential(Ref. 0093). |
|||||||||||||||||||
| 66 | 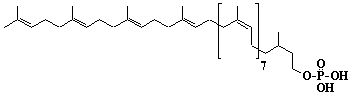 |
Dolichyl-12 monophosphate, a-Dihydrododecaprenyl monophosphate |
3,7,11,15,19,23,27,31,35,39,43,47-Dodecamethyl-6,10,14,18,22,26,30,34,38,42,46-octatetracontaundecaen-1-yl monophosphate |
IIP0066 | Yoshichika Yoshioka |
Dol-12-P |
C60H101O4P | 917.415 | The dynamics of dolichyl phosphate molecules in membranes can be regulated by transmembrane electrical potential(Ref. 0093). |
|||||||||||||||||||
| 67 | 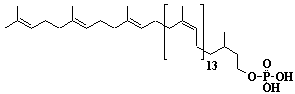 |
Dolichyl-17 monophosphate, a-Dihydroheptadecaprenyl monophosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58E,62E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67-Heptadecamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66-octahexacontahexadecaen-1-yl monophosphate |
IIP0067 | Yoshichika Yoshioka |
Dol-17-P |
C85H141O4P | 1258.000 | Role of Dol-P in regulation of protein glycosylation in 2- acetylaminofluorene- induced carcinogenesis in rat liver(Ref. 0050). Effect of Dol-P concentration on the formation of lipid-saccharides and the N-glycosylation of protein by thyroid ER vesicles(Ref. 0095). The microsomal fraction of liver has been found to catalyze glucose transfer from UDPG to a lipid acceptor to be identical to the compound obtained by chemical phosphorylation of Dol (Ref. 0096). The biological role of Dol (review) (Ref. 0043). Metabolic labeling of rat liver slices with mevalonic acid revealed the accumulation of E,E,E-geranylgeranyl (di)phosphate as well as dolichyl (di)phosphate (C85 and C90) and dehydrodolichol (C85 and C90), but no accumulation of Z,E,E-geranylgeranyl (di)phosphate or E,E-farnesyl (di)phosphate was detected(Ref. 0026). Dol-P pathway leading to N-glycosylation of protein(Ref. 0097). Content of dolichol and activity of GlcNAc-1-P (UDP-N-acetylglucosamine:dolichyl phosphate N-acetylglucosamine 1-phosphate transferase) in fibroblasts from patients with carbohydrate- deficient- glycoprotein (CDG) syndrome were analyzed. The major defect of CDG syndrome is not found in the synthesys of Dol-P or GlcNAc-PP-Dol(Ref. 0061). Dolichyl phosphate is a potent inducer of apoptosis in rat glioma cells. Apoptotic changes were also observed in HL-60 cells, SIRC cells and HeLa cells(Ref. 0098). Exogenous dolichyl phosphate induced apoptosis in the human monoblastic leukemia cell line U937 within 4 hours. Dol-P-induced apoptosis is mediated by mitogen-activated protein kinase (MAP kinase) cascade(Ref. 0099). Treatment of U937 cells with dolichyl phosphate led to an increase in the activity of the ICE family protease CPP32, accompanied with cleavage of pre-CPP32 to generate p17. The modulation of cAMP synthesis due to the stimulation of adenylate cyclase by dolichyl phosphate plays a critical role in CPP32 activation and apoptosis(Ref. 0100). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dolichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
HPLC (with chromatograph)(Ref. 0094/0048/0051). Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate in human serum were made fluorometrically by HPLC. The calibration curve of dolichol standard against the internal standard showed linearity at dolichol concentrations of 10ng to 500ng(Ref. 0053). |
Direct estimation of dolichyl phosphate in rat liver by high pressure liquid chromatography(Ref. 0094). Human liver and testis(Ref. 0048). Rat liver(Ref. 0048). Contents in human tissues (adrenal, aorta, colon, heart, kidney, liver, lung, muscle, pancreas, pituitary gland, prostate, small intestine, spleen, testis, tyroid grand, and stomach)(Ref. 0055). Rat and human tissues (spleen, liver, kidney, myocardium, lung, pancreas, adrenal)(Ref. 0051). Levels of dolichyl phosphate in neonatal and adult human tissues (liver, heart, kidney, lung, spleen)(Ref. 0056). Content in human serum(Ref. 0053). Distribution of radiolabeled dolichol-P in rat liver fractions (nuclei, debris, mitochondria, lysosomes, microsomes, Golgi, cytosol)(Ref. 0060). |
Incorporation of tritium from tritiated water into dol-P in the kidneys and liver of mice as a function of age(Ref. 0066). Age-associated changes in dol-P metabolism in the kidneys and liver of mice(Ref. 0066). Rat liver hyperplastic nodules(Ref. 0048). Human liver cirrosis and hepatocarcinoma(Ref. 0048). The amount was greatly decreased in hyperplastic nodules of rat liver(Ref. 0050). Effect of age on the content in brain, liver, spleen, kidney, and testis of the rat(Ref. 0067). Age-related chnges of dolichyl phosphate content in rat and human tissues (Ref. 0051). The uptake of F-PP and GG-PP was studied using primary cultures of chromaffin cells. Dolichylmonophosphate and isopentenylpyrophosphate promoted the uptake(Ref. 0079). Contents of various organs of mice used as a model for Niemann-Pick's type C disease . Contents in subfractions isolated from the liver of mice used as a model for NPC (Ref. 0071). The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of dolichol-P and acitivated the oligosaccharyltransferase(Ref. 0072). The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capasity were as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and dolichol were not inhibitors(Ref. 0039). Modifications in the concentration and distribution of rat liver microsomal dolichol and dolichyl phosphate after acute ethanol adiministration (6 g/kg body weight) were investigated. Between 3 and 24 hr after ethanol administration, the microsomal dolichyl phosphate concentration was significantly lower than in control rats. An earlier and more marked reduction of total dolichol was observed in the Golgi apparatus(Ref. 0059). Pro-oxidant agents lead to a significant decrease of both free dolichol and dolichyl phosphate in isolated rat hepatocytes(Ref. 0059). Distribution of radiolabeled dolichol-P in rat liver fractions(Ref. 0060). The presence of covalently bound dolichyl phosphate in rat liver(Ref. 0060). |
Monoclonal antibody to amphomycin is a tool to study the topography of dolichol monophosphate in the membrane(Ref. 0101). |
|||||||||||||||
| 68 | 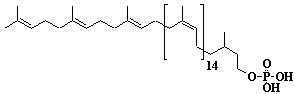 |
Dolichyl-18 monophosphate, a-Dihydrooctadecaprenyl monophosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62E,66E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71-Octadecamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70-doheptacontaheptadecaen-1-yl monophosphate |
IIP0068 | Yoshichika Yoshioka |
Dol-18-P |
C90H149O4P | 1326.117 | Role of Dol-P in regulation of protein glycosylation in 2-acetylaminofluorene-induced carcinogenesis in rat liver(Ref. 0050). Effect of Dol-P concentration on the formation of lipid-saccharides and the N-glycosylation of protein by thyroid ER vesicles(Ref. 0095). The microsomal fraction of liver has been found to catalyze glucose transfer from UDPG to a lipid acceptor to be identical to the compound obtained by chemical phosphorylation of Dol (Ref. 0096). The biological role of dolichol (review) (Ref. 0043). Metabolic labeling of rat liver slices with mevalonic acid revealed the accumulation of E,E,E-geranylgeranyl (di)phosphate as well as dolichyl (di)phosphate (C85 and C90) and dehydrodolichol (C85 and C90), but no accumulation of Z,E,E-geranylgeranyl (di)phosphate or E,E-farnesyl (di)phosphate was detected(Ref. 0026). Dol-P pathway leading to N-glycosylation of protein(Ref. 0097). Content of Dol and activity of GlcNAc-1-P (UDP-N-acetylglucosamine: Dol-P N-acetylglucosamine 1-phosphate transferase) in fibroblasts from patients with carbohydrate-deficient-glycoprotein (CDG) syndrome were analyzed. The major defect of CDG syndrome is not found in the synthesys of Dol-P or GlcNAc-PP-Dol (Ref. 0061). Dolichyl phosphate is a potent inducer of apoptosis in rat glioma cells. Apoptotic changes were also observed in HL-60 cells, SIRC cells and HeLa cells(Ref. 0098). Exogenous dolichyl phosphate induced apoptosis in the human monoblastic leukemia cell line U937 within 4 hours. Dol-P-induced apoptosis is mediated by mitogen-activated protein kinase (MAP kinase) cascade(Ref. 0099). Treatment of U937 cells with dolichyl phosphate led to an increase in the activity of the ICE family protease CPP32, accompanied with cleavage of pre-CPP32 to generate p17. The modulation od cAMP synthesis due to the stimulation of adenylate cyclase by dolichyl phosphate plays a critical role in CPP32 activation and apoptosis(Ref. 0100). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dolichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
HPLC (with chromatograph)(Ref. 0094/0048/0051). Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate in human serum were made fluorometrically by HPLC. The calibration curve of dolichol standard against the internal standard showed linearity at dolichol concentrations of 10ng to 500ng(Ref. 0053). |
Direct estimation of dolichyl phosphate in rat liver by high pressure liquid chromatography(Ref. 0094). Human liver and testis(Ref. 0048). Rat liver(Ref. 0048). Contents in human tissues (adrenal, aorta, colon, heart, kidney, liver, lung, muscle, pancreas, pituitary gland, prostate, small intestine, spleen, testis, tyroid grand, and stomach)(Ref. 0055). Rat and human tissues (spleen, liver, kidney, myocardium, lung, pancreas, adrenal)(Ref. 0051). Levels of dolichyl phosphate in neonatal and adult human tissues (liver, heart, kidney, lung, spleen)(Ref. 0056). Content in human serum(Ref. 0053). Distribution of radiolabeled dolichol-P in rat liver fractions (nuclei, debris, mitochondria, lysosomes, microsomes, Golgi, cytosol)(Ref. 0060). |
Incorporation of tritium from tritiated water into Dol-P in the kidneys and liver of mice as a function of age(Ref. 0066). Age-associated changes in Dol-P metabolism in the kidneys and liver of mice(Ref. 0066). Rat liver hyperplastic nodules(Ref. 0048). Human liver cirrosis and hepatocarcinoma(Ref. 0048). The amount was greatly decreased in hyperplastic nodules of rat liver(Ref. 0050). Effect of age on the content in brain, liver, spleen, kidney, and testis of the rat(Ref. 0067). Age-related chnges of Dol-P content in rat and human tissues (Ref. 0051). The uptake of F-PP and GG-PP was studied using primary cultures of chromaffin cells. Dolichylmonophosphate and isopentenylpyrophosphate promoted the uptake(Ref. 0079). Contents of various organs of mice used as a model for Niemann-Pick's type C disease. Contents in subfractions isolated from the liver of mice used as a model for NPC (Ref. 0071). The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of Dol-P and acitivated the oligosaccharyltransferase(Ref. 0072). The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capasity were as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and dolichol were not inhibitors(Ref. 0039). Modifications in the concentration and distribution of rat liver microsomal dolichol and dolichyl phosphate after acute ethanol adiministration (6 g/kg body weight) were investigated. Between 3 and 24 hr after ethanol administration, the microsomal dolichyl phosphate concentration was significantly lower than in control rats. An earlier and more marked reduction of total dolichol was observed in the Golgi apparatus(Ref. 0059). Pro-oxidant agents lead to a significant decrease of both free Dol and Dol-P in isolated rat hepatocytes(Ref. 0059). Distribution of radiolabeled Dol-P in rat liver fractions(Ref. 0060). The presence of covalently bound Dol-P in rat liver(Ref. 0060). |
Monoclonal antibody to amphomycin is a tool to study the topography of dolichol monophosphate in the membrane(Ref. 0101). |
|||||||||||||||
| 69 | 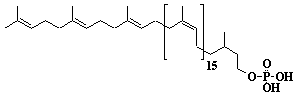 |
Dolichyl-19 monophosphate, a-Dihydrononadecaprenyl monophosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66E,70E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75-Nonadecamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74-hexaheptacontaoctadecaen-1-yl monophosphate |
IIP0069 | Yoshichika Yoshioka |
Dol-19-P |
C95H157O4P | 1394.234 | Role of dolichyl phosphate in regulation of protein glycosylation in 2-acetylaminofluorene- induced carcinogenesis in rat liver(Ref. 0050). Effect of Dol-P concentration on the formation of lipid-saccharides and the N-glycosylation of protein by thyroid ER vesicles (Ref. 0095). The microsomal fraction of liver has been found to catalyze glucose transfer from UDPG to a lipid acceptor to be identical to the compound obtained by chemical phosphorylation of Dol (Ref. 0096). The biological role of Dol (review) (Ref. 0043). Dol-P pathway leading to N-glycosylation of protein(Ref. 0097). Content of Dol and activity of GlcNAc-1-P (UDP-N-acetylglucosamine:Dol-P N-acetylglucosamine 1-phosphate transferase) in fibroblasts from patients with carbohydrate-deficient-glycoprotein (CDG) syndrome were analyzed. The major defect of CDG syndrome is not found in the synthesys of Dol-P or GlcNAc-PP-Dol (Ref. 0061). Dol-P is a potent inducer of apoptosis in rat glioma cells. Apoptotic changes were also observed in HL-60 cells, SIRC cells and HeLa cells (Ref. 0098). Exogenous dolichyl phosphate induced apoptosis in the human monoblastic leukemia cell line U937 within 4 hours. Dol-P-induced apoptosis is mediated by mitogen-activated protein kinase (MAP kinase) cascade(Ref. 0099). Treatment of U937 cells with dolichyl phosphate led to an increase in the activity of the ICE family protease CPP32, accompanied with cleavage of pre-CPP32 to generate p17. The modulation od cAMP synthesis due to the stimulation of adenylate cyclase by dolichyl phosphate plays a critical role in CPP32 activation and apoptosis(Ref. 0100). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dolichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
The head and tail sites of dolichol exibit a fast motion as exibited in the short chain isoprenols. Dolichol appears to induce a non-bilayer (isotropic) organization of phospholipid molecules in PE/PC (2:1) vesicles.(Ref. 0085) |
HPLC (with chromatograph)(Ref. 0094/0048/0051). Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate in human serum were made fluorometrically by HPLC. The calibration curve of dolichol standard against the internal standard showed linearity at dolichol concentrations of 10ng to 500ng(Ref. 0053). |
Direct estimation of dolichyl phosphate in rat liver by high pressure liquid chromatography(Ref. 0094). Human liver and testis(Ref. 0048). Rat liver(Ref. 0048). Contents in human tissues (adrenal, aorta, colon, heart, kidney, liver, lung, muscle, pancreas, pituitary gland, prostate, small intestine, spleen, testis, tyroid grand, and stomach)(Ref. 0055). Rat and human tissues (spleen, liver, kidney, myocardium, lung, pancreas, adrenal)(Ref. 0051). Levels of dolichyl phosphate in neonatal and adult human tissues (liver, heart, kidney, lung, spleen)(Ref. 0056). Content in human serum(Ref. 0053). Distribution of radiolabeled dolichol-P in rat liver fractions (nuclei, debris, mitochondria, lysosomes, microsomes, Golgi, cytosol)(Ref. 0060). |
Incorporation of tritium from tritiated water into Dol-P in the kidneys and liver of mice as a function of age(Ref. 0066). Age-associated changes in Dol-P metabolism in the kidneys and liver of mice(Ref. 0066). Rat liver hyperplastic nodules(Ref. 0048). Human liver cirrosis and hepatocarcinoma(Ref. 0048). The amount was greatly decreased in hyperplastic nodules of rat liver(Ref. 0050). Effect of age on the content in brain, liver, spleen, kidney, and testis of the rat(Ref. 0067). Age-related chnges of Dol-P content in rat and human tissues (Ref. 0051). The uptake of F-PP and GG-PP was studied using primary cultures of chromaffin cells. Dol-P and isopentenylpyrophosphate promoted the uptake(Ref. 0079). Contents of various organs of mice used as a model for Niemann-Pick's type C disease (liver, spleen, kidney, brain, testis, lung, adipose tissue, heart, intestine, skeletal muscle). Contents in subfractions isolated from the liver of mice used as a model for NPC (mitocondria, lysosomes, microsomes) (Ref. 0071). The effects of TSH on Dol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of Dol-P and acitivated the oligosaccharyltransferase (Ref. 0072). The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capasity were as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and Dol were not inhibitors(Ref. 0039). Modifications in the concentration and distribution of rat liver microsomal Dol and Dol-Pafter acute ethanol adiministration (6 g/kg body weight) were investigated. Between 3 and 24 hr after ethanol administration, the microsomal Dol-P concentration was significantly lower than in control rats. An earlier and more marked reduction of total Dol was observed in the Golgi apparatus(Ref. 0059). Pro-oxidant agents lead to a significant decrease of both freeDol and Dol-P in isolated rat hepatocytes(Ref. 0059). Distribution of radiolabeled Dol-P in rat liver fractions(Ref. 0060). The presence of covalently bound Dol-P in rat liver(Ref. 0060). |
Monoclonal antibody to amphomycin is a tool to study the topography of dolichol monophosphate in the membrane(Ref. 0101). |
||||||||||||||
| 70 | 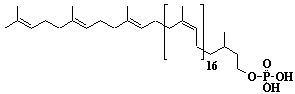 |
Dolichyl-20 monophosphate, a-Dihydroeicosaprenyl monophosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66Z,70E,74E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75,79-Eicosamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74,78-octacontanonadecaen-1-yl monophosphate |
IIP0070 | Yoshichika Yoshioka |
Dol-20-P |
C100H165O4P | 1462.351 | Role of Dol-P in regulation of protein glycosylation in 2-acetylaminofluorene-induced carcinogenesis in rat liver(Ref. 0050). Effect of Dol-P concentration on the formation of lipid-saccharides and the N-glycosylation of protein by thyroid ER vesicles(Ref. 0095). The microsomal fraction of liver has been found to catalyze glucose transfer from UDPG to a lipid acceptor to be identical to the compound obtained by chemical phosphorylation of Dol (Ref. 0096). The biological role of Dol (review) (Ref. 0043). Dol-P pathway leading to N-glycosylation of protein(Ref. 0097). Content of Dol and activity of GlcNAc-1-P (UDP-N-acetylglucosamine:Dol-P N-acetylglucosamine 1-phosphate transferase) in fibroblasts from patients with carbohydrate-deficient-glycoprotein (CDG) syndrome were analyzed. The major defect of CDG syndrome is not found in the synthesys of Dol-P or GlcNAc-PP-Dol(Ref. 0061). Dol-P is a potent inducer of apoptosis in rat glioma cells. Apoptotic changes were also observed in HL-60 cells, SIRC cells and HeLa cells(Ref. 0098). Exogenous Dol-P induced apoptosis in the human monoblastic leukemia cell line U937 within 4 hours. Dol-P-induced apoptosis is mediated by mitogen-activated protein kinase (MAP kinase) cascade(Ref. 0099). Treatment of U937 cells with Dol-P led to an increase in the activity of the ICE family protease CPP32, accompanied with cleavage of pre-CPP32 to generate p17. The modulation od cAMP synthesis due to the stimulation of adenylate cyclase by Dol-P plays a critical role in CPP32 activation and apoptosis(Ref. 0100). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dolichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
HPLC (with chromatograph)(Ref. 0094/0048/0051). Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate in human serum were made fluorometrically by HPLC. The calibration curve of dolichol standard against the internal standard showed linearity at dolichol concentrations of 10ng to 500ng(Ref. 0053). |
Direct estimation of dolichyl phosphate in rat liver by high pressure liquid chromatography(Ref. 0094). Human liver and testis(Ref. 0048). Rat liver(Ref. 0048). Contents in human tissues (adrenal, aorta, colon, heart, kidney, liver, lung, muscle, pancreas, pituitary gland, prostate, small intestine, spleen, testis, tyroid grand, and stomach)(Ref. 0055). Rat and human tissues (spleen, liver, kidney, myocardium, lung, pancreas, adrenal)(Ref. 0051). Levels of dolichyl phosphate in neonatal and adult human tissues (liver, heart, kidney, lung, spleen)(Ref. 0056). Content in human serum(Ref. 0053). Distribution of radiolabeled dolichol-P in rat liver fractions (nuclei, debris, mitochondria, lysosomes, microsomes, Golgi, cytosol)(Ref. 0060). |
Phosphorylation of dolichol-20. |
Incorporation of tritium from tritiated water into Dol-P in the kidneys and liver of mice as a function of age(Ref. 0066). Age-associated changes in Dol-P metabolism in the kidneys and liver of mice(Ref. 0066). Rat liver hyperplastic nodules(Ref. 0048). Human liver cirrosis and hepatocarcinoma(Ref. 0048). The amount was greatly decreased in hyperplastic nodules of rat liver(Ref. 0050). Effect of age on the content in brain, liver, spleen, kidney, and testis of the rat(Ref. 0067). Age-related chnges of Dol-P content in rat and human tissues (spleen, liver, kidney, heart, lung, pancreas, adrenal)(Ref. 0051). The uptake of F-PP and GG-PP was studied using primary cultures of chromaffin cells. Dolichylmonophosphate and isopentenylpyrophosphate promoted the uptake(Ref. 0079). Contents of various organs of mice used as a model for Niemann-Pick's type C disease (liver, spleen, kidney, brain, testis, lung, adipose tissue, heart, intestine, skeletal muscle). Contents in subfractions isolated from the liver of mice used as a model for NPC (mitocondria, lysosomes, microsomes)(Ref. 0071). The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of Dol-P and acitivated the oligosaccharyltransferase (Ref. 0072). The potential for feedback inhibition by isoprene intermediates in the isoprene biosynthetic pathway was investigated. The relative inhibitory capasity were as follows: GG-PP > F-PP > G-PP > FOH > dol-P. GOH and Dol were not inhibitors(Ref. 0039). Modifications in the concentration and distribution of rat liver microsomal Dol and Dol-P after acute ethanol adiministration (6 g/kg body weight) were investigated. Between 3 and 24 hr after ethanol administration, the microsomal Dol-P concentration was significantly lower than in control rats. An earlier and more marked reduction of total Dol was observed in the Golgi apparatus(Ref. 0059). Pro-oxidant agents lead to a significant decrease of both free Dol and Dol-P in isolated rat hepatocytes(Ref. 0059). Distribution of radiolabeled Dol-P in rat liver fractions(Ref. 0060). The presence of covalently bound Dol-P in rat liver(Ref. 0060). |
The effect of dolichols, polyprenols, dolichol esterified with fatty acids, and dolichyl phosphate on the structure and fluidity of model membranes was studied using 31P NMR, small-angle x-ray scattering, differential scanning calorimetry, and freeze-fracture electron microscopy. Dolichol and dolichol derivatives destabilize unsaturated phosphatidylethanolamine containing bilayer structures and promote hexagonal II phase formation. Dolichol and dolichyl-P increase the fatty acid fluidity in phosphatidylethanolamine mixtures(Ref. 0044). Monoclonal antibody to amphomycin is a tool to study the topography of dolichol monophosphate in the membrane(Ref. 0101). |
||||||||||||||
| 71 | 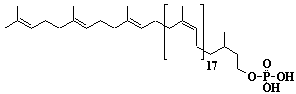 |
Dolichyl-21 monophosphate, a-Dihydroheneicosaprenyl monophosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66Z,70Z,74E,78E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75,79,83-Henicosamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74,78,82-tetraoctacontaeicosaen-1-yl monophosphate |
IIP0071 | Yoshichika Yoshioka |
Dol-21-P |
C105H173O4P | 1530.468 | Role of Dol-P in regulation of protein glycosylation in 2-acetylaminofluorene-induced carcinogenesis in rat liver(Ref. 0050). Effect of Dol-P concentration on the formation of lipid-saccharides and the N-glycosylation of protein by thyroid ER vesicles(Ref. 0095). The microsomal fraction of liver has been found to catalyze glucose transfer from UDPG to a lipid acceptor to be identical to the compound obtained by chemical phosphorylation of Dol(Ref. 0096). The biological role of Dol (review)(Ref. 0043). Dol-P pathway leading to N-glycosylation of protein(Ref. 0097). Content of Dol and activity of GlcNAc-1-P (UDP-N-acetylglucosamine:Dol-P N-acetylglucosamine 1-phosphate transferase) in fibroblasts from patients with carbohydrate-deficient-glycoprotein (CDG) syndrome were analyzed. The major defect of CDG syndrome is not found in the synthesys of dolichyl phosphate or GlcNAc-PP-Dol(Ref. 0061). Dol-P is a potent inducer of apoptosis in rat glioma cells. Apoptotic changes were also observed in HL-60 cells, SIRC cells and HeLa cells(Ref. 0098). Exogenous Dol-P induced apoptosis in the human monoblastic leukemia cell line U937 within 4 hours. Dol-P-induced apoptosis is mediated by mitogen-activated protein kinase (MAP kinase) cascade(Ref. 0099). Treatment of U937 cells with Dol-P led to an increase in the activity of the ICE family protease CPP32, accompanied with cleavage of pre-CPP32 to generate p17. The modulation od cAMP synthesis due to the stimulation of adenylate cyclase by Dol-P plays a critical role in CPP32 activation and apoptosis(Ref. 0100). Dolichyl, dihydroheptaprenyl, and dihydrodecaprenyl phosphates induced apoptosis in U937 cells, while farnesyl phosphate, geranylgeranyl phosphate, dolichol, dolichal, and dolichoic acid did not induce apoptosis. Apoptosis induced by dihydroprenyl phosphate was mediated by caspase-3-like activation but not by caspase-1-like activation(Ref. 0065). |
HPLC (with chromatograph)(Ref. 0094/0048/0051). Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate in human serum were made fluorometrically by HPLC. The calibration curve of dolichol standard against the internal standard showed linearity at dolichol concentrations of 10ng to 500ng(Ref. 0053). |
Direct estimation of dolichyl phosphate in rat liver by high pressure liquid chromatography(Ref. 0094). Human liver and testis(Ref. 0048). Rat liver(Ref. 0048). Contents in human tissues (adrenal, aorta, colon, heart, kidney, liver, lung, muscle, pancreas, pituitary gland, prostate, small intestine, spleen, testis, tyroid grand, and stomach)(Ref. 0055). Rat and human tissues (spleen, liver, kidney, myocardium, lung, pancreas, adrenal)(Ref. 0051). Levels of dolichyl phosphate in neonatal and adult human tissues (liver, heart, kidney, lung, spleen)(Ref. 0056). Content in human serum(Ref. 0053). |
Rat liver hyperplastic nodules(Ref. 0009). Human liver cirrosis and hepatocarcinoma (Ref. 0048). The amount was greatly decreased in hyperplastic nodules of rat liver(Ref. 0050). Effect of age on the content in brain, liver, spleen, kidney, and testis of the rat(Ref. 0067). Age-related chnges of Dol-P content in rat and human tissues (spleen, liver, kidney, heart, lung, pancreas, adrenal)(Ref. 0051). The uptake of F-PP and GG-PP was studied using primary cultures of chromaffin cells. Dolichylmonophosphate and isopentenyl pyrophosphate promoted the uptake (Ref. 0079). Contents of various organs of mice used as a model for Niemann-Pick's type C disease (liver, spleen, kidney, brain, testis, lung, adipose tissue, heart, intestine, skeletal muscle). Contents in subfractions isolated from the liver of mice used as a model for NPC (mitocondria, lysosomes, microsomes)(Ref. 0071). The effects of TSH on Dol-linked sugar pools and oligosaccharyl transferase activity in thyroid cells were investigated. TSH slightly increased the amount of Dol-P and acitivated the oligosaccharyltransferase(Ref. 0072). Modifications in the concentration and distribution of rat liver microsomal dolichol and dolichyl phosphate after acute ethanol adiministration (6 g/kg body weight) were investigated. Between 3 and 24 hr after ethanol administration, the microsomal dolichyl phosphate concentration was significantly lower than in control rats. An earlier and more marked reduction of total Dol was observed in the Golgi apparatus(Ref. 0059). Pro-oxidant agents lead to a significant decrease of both free Dol and Dol-P in isolated rat hepatocytes(Ref. 0059). |
Monoclonal antibody to amphomycin is a tool to study the topography of dolichol monophosphate in the membrane(Ref. 0101). |
|||||||||||||||
| 72 | 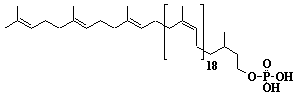 |
Dolichyl-22 monophosphate, a-Dihydrodocosaprenyl monophosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66Z,70Z,74Z,78E,82E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75,79,83,87-Docosamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74,78,82,86-octaoctacontahenicosaen-1-yl monophosphate |
IIP0072 | Yoshichika Yoshioka |
Dol-22-P |
C110H181O4P | 1598.586 | The biological role of dolichol (review)(Ref. 0043). Dolichyl phosphate is a potent inducer of apoptosis in rat glioma cells. Apoptotic changes were also observed in HL-60 cells, SIRC cells and HeLa cells(Ref. 0098). Treatment of U937 cells with dolichyl phosphate led to an increase in the activity of the ICE family protease CPP32, accompanied with cleavage of pre-CPP32 to generate p17. The modulation od cAMP synthesis due to the stimulation of adenylate cyclase by dolichyl phosphate plays a critical role in CPP32 activation and apoptosis(Ref. 0100). |
Rat liver hyperplastic nodules(Ref. 0048). Human liver cirrosis and hepatocarcinoma(Ref. 0048). Age-related chnges of dolichyl phosphate content in rat and human tissues (spleen, liver, kidney, heart, lung, pancreas, adrenal)(Ref. 0051). Contents of various organs of mice used as a model for Niemann-Pick's type C disease (liver, spleen, kidney, brain, testis, lung, adipose tissue, heart, intestine, skeletal muscle). Contents in subfractions isolated from the liver of mice used as a model for NPC (mitocondria, lysosomes, microsomes)(Ref. 0071). |
||||||||||||||||||
| 73 | 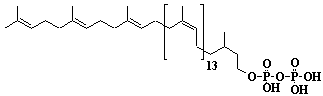 |
Dolichyl-17 diphosphate, a-Dihydroheptadecaprenyl diphosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58E,62E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67-Heptadecamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66-octahexacontahexadecaen-1-yl diphosphate |
IIP0073 | Yoshichika Yoshioka |
Dol-17-PP |
C85H142O7P2 | 1337.980 | Calf brain microsomes catalyze the enzymatic release of P from exogenous Dol-PP by a bacitracin-sensitive reaction(Ref. 0102). Metabolic labeling of rat liver slices with mevalonic acid revealed the accumulation of E,E,E-geranylgeranyl (di)phosphate as well as dolichyl (di)phosphate (C85 and C90) and dehydrodolichol (C85 and C90), but no accumulation of Z,E,E-geranylgeranyl (di)phosphate or E,E-farnesyl (di)phosphate was detected(Ref. 0026). |
The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of dolichol-PP and acitivated the oligosaccharyltransferase(Ref. 0072). |
||||||||||||||||||
| 74 | 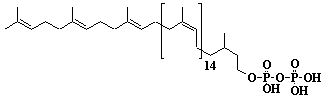 |
Dolichyl-18 diphosphate, a-Dihydrooctadecaprenyl diphosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62E,66E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71-Octadecamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70-doheptacontaheptadecaen-1-yl diphosphate |
IIP0074 | Yoshichika Yoshioka |
Dol-18-PP |
C90H150O7P2 | 1406.097 | Metabolic labeling of rat liver slices with mevalonic acid revealed the accumulation of E,E,E-geranylgeranyl (di)phosphate as well as dolichyl (di)phosphate (C85 and C90) and dehydrodolichol (C85 and C90), but no accumulation of Z,E,E-geranylgeranyl (di)phosphate or E,E-farnesyl (di)phosphate was detected(Ref. 0026). |
The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of dolichol-PP and acitivated the oligosaccharyltransferase(Ref. 0072). |
||||||||||||||||||
| 75 | 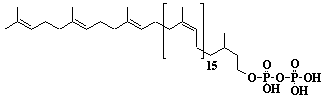 |
Dolichyl-19 diphosphate, a-Dihydrononadecaprenyl diphosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66E,70E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75-Nonadecamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74-hexaheptacontaoctadecaen-1-yl diphosphate |
IIP0075 | Yoshichika Yoshioka |
Dol-19-PP |
C95H158O7P2 | 1474.214 | The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of dolichol-PP and acitivated the oligosaccharyltransferase(Ref. 0072). |
|||||||||||||||||||
| 76 | 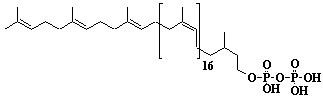 |
Dolichyl-20 diphosphate, a-Dihydroeicosaprenyl diphosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66Z,70E,74E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75,79-Eicosamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74,78-octacontanonadecaen-1-yl diphosphate |
IIP0076 | Yoshichika Yoshioka |
Dol-20-PP |
C100H166O7P2 | 1542.331 | The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of dolichol-PP and acitivated the oligosaccharyltransferase(Ref. 0072). |
|||||||||||||||||||
| 77 | 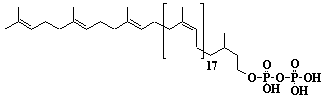 |
Dolichyl-21 diphosphate, a-Dihydroheneicosaprenyl diphosphate |
[6Z,10Z,14Z,18Z,22Z,26Z,30Z,34Z,38Z,42Z,46Z,50Z,54Z,58Z,62Z,66Z,70Z,74E,78E]-3,7,11,15,19,23,27,31,35,39,43,47,51,55,59,63,67,71,75,79,83-Henicosamethyl-6,10,14,18,22,26,30,34,38,42,46,50,54,58,62,66,70,74,78,82-tetraoctacontaeicosaen-1-yl diphosphate |
IIP0077 | Yoshichika Yoshioka |
Dol-21-PP |
C105H174O7P2 | 1610.448 | The effects of TSH on dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells were investigated. TSH slightly increased the amount of dolichol-PP and acitivated the oligosaccharyltransferase(Ref. 0072). |
|||||||||||||||||||
| 78 |  |
C12 a, w-dibasic prenoic acid |
[2E,6E]-3,7-Dimethyl-2,6-decadien-1,10-dioic acid |
IIP0078 | Hiroshi Sagami |
C12H18O4 | 226.269 | Isolated embryonic Drosophila cell extracts metabolize farnesyl diphosphate and farnesol to identical products. Two of the major products were identified as trans,trans-3,7,11-trimethyl-2,6,10-dodecatrien-1,12-dioic acid and trans-3,7-dimethyl-2,6-decadien-1,10-dioic acid.(Ref. 0103) |
||||||||||||||||||||
| 79 |  |
C15 a, w-dibasic prenoic acid |
[2E,6E,10E]-3,7,11-Trimethyl-2,6,10-dodecatrien-1,12-dioic acid |
IIP0079 | Hiroshi Sagami |
C15H22O4 | 266.333 | Isolated embryonic Drosophila cell extracts metabolize farnesyl diphosphate and farnesol to identical products. Two of the major products were identified as trans,trans-3,7,11-trimethyl-2,6,10-dodecatrien-1,12-dioic acid and trans-3,7-dimethyl-2,6-decadien-1,10-dioic acid.(Ref. 0103) |
||||||||||||||||||||
| 80 |  |
All-trans Farnesyl amine |
[2E,6E]-3,7,11-Trimethyl-2,6,10-dodecatrienyl amine |
IIP0080 | Hiroshi Sagami |
F-NH2 |
C15H27N | 221.382 | Farnesylamine, an analogue of farnesol, inhibit growth of PAP2 cells in a dose-dependent manner (Ref. 0104). |
All-trans farnesylamine can be synthesized from all-trans farnesol by the Gabriel synthesis (Ref. 0104). |
||||||||||||||||||
| 81 | 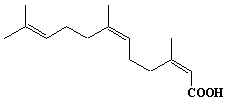 |
cis,cis-Farnesol |
[2Z,6Z]-3,7,11-Trimethyl-2,6,10-dodecatrienic acid |
IIP0081 | Hiroshi Sagami |
C15H24O2 | 236.350 | cis,cis-Farnesoic acid , trans,trans-farnesoic acid, trans,trans,trans-geranylgeranoic acid, and an unidentified C20 isoprenoid acid containing at least one cis-double bond was found as mevalonate-derived metabolites with cell-free homogenates of bovine retinas (Ref. 0105) |
||||||||||||||||||||
| 82 | 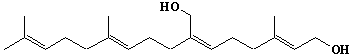 |
Plaunotol |
[2E,6Z,10E]-7-Hydroxymethyl-3,11,15-trimethyl-2,6,10,14-hexadecatetraen-1-ol |
IIP0082 | Hiroshi Sagami |
C20H34O2 | 306.483 | Plaunotol was isolated as a potent antipeptic ulcer substance from a Thai medicinal plant identified with Croton sublyratus Kurz.(Ref. 0107) |
||||||||||||||||||||
| 83 | 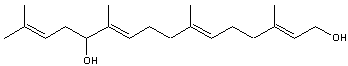 |
12(S)-hydroxygeranylgeraniol |
[2E,6E,10E]-12-Hydroxy-3,7,11,15-tetramethyl-2,6,10,14-hexadecatetraen-1-ol |
IIP0083 | Hiroshi Sagami |
C20H34O2 | 306.483 | Cytotoxicity against cultured human tumor cell lines, A549, SK-OV-3, SK-MEL-2, XF498, and HCT15, with ED50 values ranging from 4.1-8.3 micrograms/ml. (Ref. 0108) |
12(S)-hydroxygeranylgeraniol and (2E,10E]-1,12-dihydroxy-18-acetoxy-3,7,15-trimethylhexadeca-2,10,14-triene were isolated from the aerical parts of Carpesium divaricatum.(Ref. 0108) |
|||||||||||||||||||
| 84 | 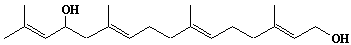 |
13(S)-hydroxygeranylgeraniol |
[2E,6E,10E]-13-Hydroxy-3,7,11,15-tetramethyl-2,6,10,14-hexadecatetraen-1-ol |
IIP0084 | Hiroshi Sagami |
C20H34O2 | 306.483 | 13(S)-Hydroxygeranylgeraniol-derived furanoditerpenes were isolated from a sample of the brown alga Bifurcaria bifurcata collected from Brittany on the Atlanticcoast.(Ref. 0109) |
||||||||||||||||||||
| 85 | 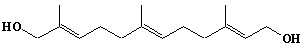 |
12-Hydroxyfarnesol |
[2E,6E,10E]-12-Hydroxy-3,7,11-trimethyl-2,6,10-dodecatrien-1-ol |
IIP0085 | Hiroshi Sagami |
C15H26O2 | 238.366 | 12-Hydroxy farnesol is formed from farnesol by the action of a cytochrome P450 terpenoid hydroxylase. (Ref. 0110) |
||||||||||||||||||||
| 86 |  |
10-Hydroxygeraniol |
[2E,6Z]-7-Hydroxymethyl-3-methyl-2,6-octadiene-1-ol |
IIP0086 | Hiroshi Sagami |
C10H18O2 | 170.249 | 10-Hydroxygeraniol is formed from geraniol by the action of acyclic monoterpene primary alcohol:NADP+ oxidereductase.(Ref. 0111) |
||||||||||||||||||||
| 87 | 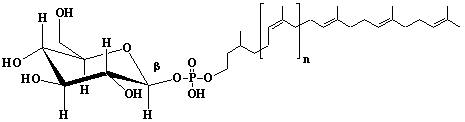 |
Glc-phosphoryldolichol (family), Glucosylphosphoryldolichol (family) |
IIP0087 | Hiroshi Sagami |
Glc-P-Dol |
|||||||||||||||||||||||
| 88 | 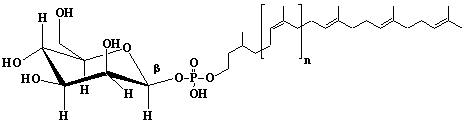 |
Man-phosphoryldolichol (family), Mannosylphosphoryldolichol (family) |
IIP0088 | Hiroshi Sagami |
Man-P-Dol |
|||||||||||||||||||||||
| 89 | 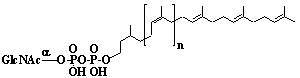 |
GlcNAc-pyrophosphoryldolichol (family), N-Acetylglcosaminylpyrophosphoryldolichol (family) |
IIP0089 | Hiroshi Sagami |
GlcNAc-PP-Dol |
|||||||||||||||||||||||
| 90 | 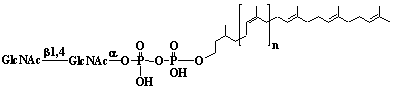 |
(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0090 | Hiroshi Sagami |
(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 91 | 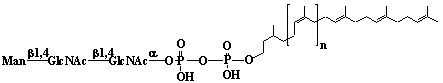 |
Man(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0091 | Hiroshi Sagami |
Man(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 92 | 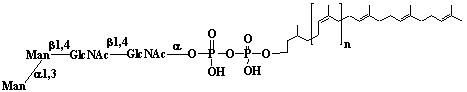 |
(Man)2(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0092 | Hiroshi Sagami |
(Man)2(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 93 | 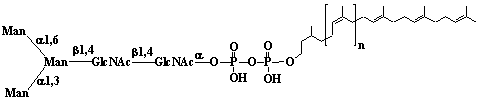 |
(Man)3(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0093 | Hiroshi Sagami |
(Man)3(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 94 | 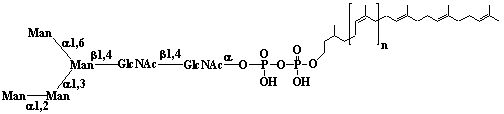 |
(Man)4(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0094 | Hiroshi Sagami |
(Man)4(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 95 | 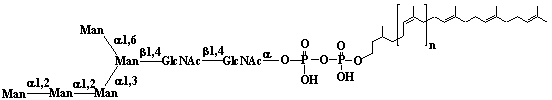 |
(Man)5(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0095 | Hiroshi Sagami |
(Man)5(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 96 | 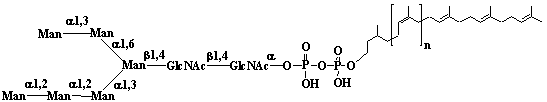 |
(Man)6(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0096 | Hiroshi Sagami |
(Man)6(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 97 | 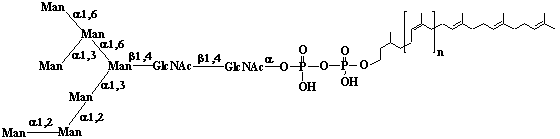 |
(Man)7(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0097 | Hiroshi Sagami |
(Man)7(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 98 | 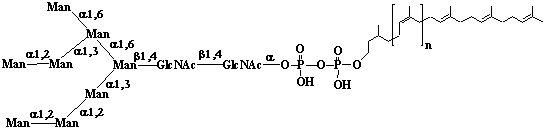 |
(Man)8(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0098 | Hiroshi Sagami |
(Man)8(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 99 | 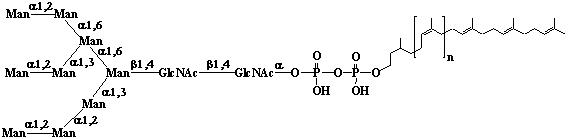 |
(Man)9(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0099 | Hiroshi Sagami |
(Man)9(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 100 | 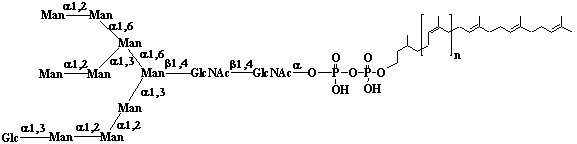 |
Glc(Man)9(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0100 | Hiroshi Sagami |
Glc(Man)9(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 101 | 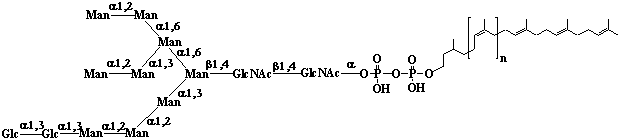 |
(Glc)2(Man)9(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0101 | Hiroshi Sagami |
(Glc)2(Man)9(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 102 | 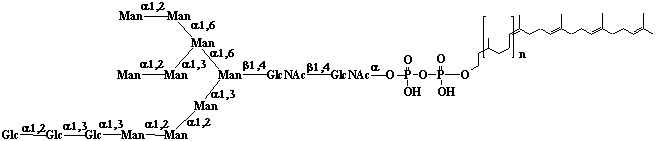 |
(Glc)3(Man)9(GlcNAc)2-pyrophosphoryldolichol (family) |
IIP0102 | Hiroshi Sagami |
(Glc)3(Man)9(GlcNAc)2-PP-Dol |
|||||||||||||||||||||||
| 103 | 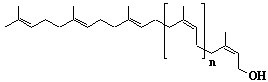 |
Dehydrodolichol (family) |
IIP0103 | Yoshichika Yoshioka |
De-dol |
Metabolic labeling of rat liver slices with mevalonic acid revealed the accumulation of E,E,E-geranylgeranyl (di)phosphate as well as dolichyl (di)phosphate (C85 and C90) and dehydrodolichol (C85 and C90), but no accumulation of Z,E,E-geranylgeranyl (di)phosphate or E,E-farnesyl (di)phosphate was detected(Ref. 0026). |
Separation of dolichol from dehydrodolichol (C75-C100) by a simple two-plate thin-layer chromatography(Ref. 0052). |
Formation of dolichol from dehydrodolichol (C90-C100) is catalyzed by NADPH- dependent reductase localized in microsomes of rat liver(Ref. 0092). Dehydrodolichol (C65-C95) was detected in the nonpolar products formed from isopentenyl diphosphate and farnesyl diphosphate in the reaction with a crude 1000 x g supernatant of yeast homogenates in the presence of NADPH. Dehydrodolichol is a common branch point intermediate in the biosynthetic pathways leading to dolichal and dolichol. Dehydrodolichal is an intermediate in the pathway from dehydrodolichol to dolichal (Ref. 0073). |
||||||||||||||||||||
| 104 | 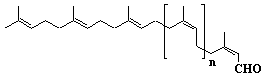 |
Dehydrodolichal (family) |
IIP0104 | Yoshichika Yoshioka |
De-dolichal |
Synthesized by treating dehydrodolichol with MnO2(Ref. 0073). |
Dehydrodolichal (C75-C80) was detected in the nonpolar products formed from isopentenyl diphosphate and farnesyl diphosphate in the reaction with a crude 1000 x g supernatant of yeast homogenates in the presence of NADPH. Dehydrodolichol is a common branch point intermediate in the biosynthetic pathways leading to dolichal and dolichol. Dehydrodolichal is an intermediate in the pathway from dehydrodolichol to dolichal (Ref. 0073). |
|||||||||||||||||||||
| 105 | 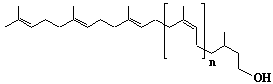 |
Dolichol (family) |
IIP0105 | Hiroshi Sagami |
Dol |
Review(Ref. 0115) |
||||||||||||||||||||||
| 106 | 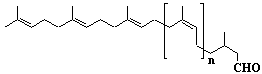 |
Dolichal (family) |
IIP0106 | Yoshichika Yoshioka |
Synthesized by treatment of dolichol with pyridinium dichromate(Ref. 0073). |
|||||||||||||||||||||||
| 107 | 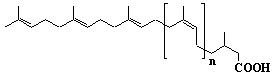 |
Dolichoic acid (family) |
IIP0107 | Yoshichika Yoshioka |
Metabolism of dolichol, dolichoic acid, and nordolichoic acid in MDCK and HepG2 cells were investigated. Dolichoic acid was broken down by alpha-oxidation and also converted to a mixture of polar compounds. No oxidative breakdown of dolichol was found(Ref. 0074). |
|||||||||||||||||||||||
| 108 | 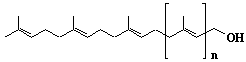 |
trans-polyprenol (family) |
IIP0108 | Hiroshi Sagami |
Chicle resin contains trans- and cis-polyisoprenes(Ref. 0116). |
Review (Structure and biosynthesis mechanism of natural polyisoprene )(Ref. 0116) |
||||||||||||||||||||||
| 109 | 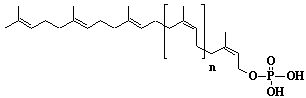 |
Dehydrodolichyl phosphate (family) |
IIP0109 | Yoshichika Yoshioka |
De-dol-P |
|||||||||||||||||||||||
| 110 | 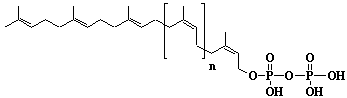 |
Dehydrodolichyl diphosphate (family) |
IIP0110 | Yoshichika Yoshioka |
De-dol-PP |
Several detergents activated microsomal dehydrodolichyl diphosphate synthase of rat liver, but the chain length of products shifted downward from C90 and C95 with increasing concentration of the detergents(Ref. 0117). |
||||||||||||||||||||||
| 111 |  |
Dolichyldolichoate (family) |
IIP0111 | Hiroshi Sagami |
Dolichyl dolichoate was found in the neutral lipid fraction of bovine thyroid(Ref. 0118). |
|||||||||||||||||||||||
| 112 | No image | Tetrahydroxybacteriohopane/bacteriohopanetetrol |
1,2,3,4-Tetrahydroxypentane-29-hopane |
IIP0112 | Yasutaka Tahara |
THBH |
C35H62O4 | 546.864 | THBH may be localized in membranes , playing there the same role as sterols play in eukaryotes , as a result of their similar size , rigidity , and amphiphilic character . (Ref. 0005) |
181-182 |
Solubled in chloroform |
3340,2920,2860,1360,1140,1110cm-1(Ref. 0001) |
1H-NMR(CDCl3):d0.683(3H,s,18a-CH3),0.791(3h,s,4b-CH3),0.812(3h,s,4a-CH3),0.846(3h,s,10b-CH3),0.904(3h,d,J=6,22R-CH3),0.944(6H,s,8band 14aCH3),2.055(3H,s,CH3COO-),2.076(3h,s,CH3COO-),2.083(3H,s,CH3COO-)2.088(3H,s,CH3COO-),4.15(1H,dd,J35a,35=12,J35a,34=6,35Ha),4.39(1H,dd,J35a,35b=12,J35a,34=3,35-Hb), 5.03(1H,dt,J3233=9,J3231=3.5,32-H), 5.24(2H,m,33-H and 34-H)(Ref. 0002)13C-NMR(CDCl3):40.4(C-1),18.7(C-2),42.1(C-3),33.4(C-4),56.2(C-5),18.7(C-6),33.3(C-7),41.8(C-8),50.5(C-9),37.5(C-10),20.9(C-11),24.0(C-12),49.4(C-13),41.8(C-14),33.8(C-15),22.8(C-16),54.5(C-17),44.4(C-18),41.6(C-19),27.5(C-20),46.0(C-21),36.1(C-22),33.4(C-23),21.6(C-24),15.9(C-25),16.6(C-26),16.6(C-27),15.9(C-28),19.8(C-29),30.9(C-30),26.3(C-31),69.6(C-32),71.7(C-33),72.0(C-34),62.1(C-35),20.6(2,20.8,20.9(CH3CO-),169.6,169.8,170.2,170.5(CH3CO-)(Ref. 0002) |
Cyanobasteria(Anabena variabilis,Calothrix sp.,Fischerella sp.,Nostoc muscorum B,Ascillatoria sp.,Scytonema sp.,Synechocystis sp.),Purple non-sulphur bacteria(Rhodomicrobium vannielii,Rhodoseudomonas acidphila,Rhodoseudomonas palustris,Rhodospirillum rubrum),Methylotrophs(Methylomonas albus,Methlomonas methanica,Methylocystis parvus,Methylosinus sporium,Methylosinus trichosporium,Methylococcus capsulatus),Gram-nagative chomoautotrophs(Nitrsomonas europaea),Gram-nagative chemoheterotrophs(Metylobacterium organophilum,Hyphomicrobiumsp.,Pseudomonas cepacia, Azotobacter chroococcum,Azotobacter vinelandii,Zymomonas mobilis),Acetic acid bacteria(A.aceti,A.liquefaciens,A.xylininum,A.pasteurianus,A.lovaniensis,A.orleanensis,A.pasteurianus,A.peroxydans,Gluconobacter oxydans),Gram-positive bacteria(Bacillus acidocaldarius,Streptomyces chartreusis,Streptomyces sp.)(Ref. 0004) |
Gene of squalene cyclase catalyzing cyclization of squalene to form hopane-22(29)-ene has been cloned and sequenced . (Ref. 0008) |
|||||||||||||
| AUTHOR | : | Toyoda, M., Asahina,M., Fukawa,H., Shimizu,T. |
| TITLE | : | Isolation of new acyclic C25-isoprenyl alcohol from potato leaves |
| JOURNAL | : | Tetrahedron Lett. |
| VOL | : | 55 PAGE : 4879-4882 (1969) |
| AUTHOR | : | Suga, T., Ohta,S., Nakai,A., and Munesada,K. |
| TITLE | : | Glycinoprenols:Novel polyprenols possesing a phytyl residue from the leaves of soybean |
| JOURNAL | : | J. Org. Chem. |
| VOL | : | 54 PAGE : 3390-3393 (1989) |
| AUTHOR | : | Fukawa,H., Toyoda, M., Shimizu, T.,and Murohashi, M. |
| TITLE | : | Isolation of new isoprenyl alcohols from silkworm feces |
| JOURNAL | : | Tetrahedron Lett. |
| VOL | : | 49 PAGE : 6209-6213 (1966) |
| AUTHOR | : | Toyoda,M.,Fukawa,H., and Shimizu, T. |
| TITLE | : | Isolation of newpolyisoprenyl ketone from silworm feces |
| JOURNAL | : | Tetrahedron Lett. |
| VOL | : | 35 PAGE : 3837-3837 (1968) |
| AUTHOR | : | Enzell, C. R., Kimland, B., and Gunnarsson, L.E. |
| TITLE | : | Tabacco Chemistry 5: Nor-solanesene, a C44-isoprenoid hydrocarbon from tabaco |
| JOURNAL | : | Tetrahedron Lett. |
| VOL | : | 22 PAGE : 1983-1986 (1971) |
| AUTHOR | : | Gough, D. P., and Hemming, F. W. |
| TITLE | : | The stereochemistry of betulaprenol biosynthesis PubMed ID:4315982 |
| JOURNAL | : | Biochem J. |
| VOL | : | 117 PAGE : 309-317 (1970) |
| AUTHOR | : | LIndgren, B. O. |
| TITLE | : | Homologous aliphatic C30-C45 terpenols in birch wood. |
| JOURNAL | : | Acta Chem. Scand. |
| VOL | : | 19 PAGE : 1317-1326 (1965) |
| AUTHOR | : | Barr, R. M., and Hemming, F. W. |
| TITLE | : | Polyprenols of Aspergillus niger. Their characterization, biosynthesis and subcellular distribution PubMed ID:5073731 |
| JOURNAL | : | Biochem J. |
| VOL | : | 126 PAGE : 1193-1202 (1972) |
| AUTHOR | : | Pennock, J. F., Hemming, F. W., and Morton, R.A. |
| TITLE | : | Dolichol: a naturally occurring isoprenoid alcohol. |
| JOURNAL | : | Nature |
| VOL | : | 186 PAGE : 470-472 (1960) |
| AUTHOR | : | Anderson, M. S., Muehlbacher, M., Street, I. P., Proffitt, J., and Poulter, C. D. |
| TITLE | : | Isopentenyl diphosphate:dimethylallyl diphosphate isomerase. An improved purification of the enzyme and isolation of the gene from Saccharomyces cerevisiae PubMed ID:2681212 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 264 PAGE : 19169-19175 (1989) |
| AUTHOR | : | Jiang, Y., Proteau, P., Poulter, D., and Ferro-Novick, S. |
| TITLE | : | BTS1 encodes a geranylgeranyl diphosphate synthase in Saccharomyces cerevisiae PubMed ID:7665600 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 270 PAGE : 21793-21799 (1995) |
| AUTHOR | : | Okada, K., Kamiya, Y., Zhu, X., Suzuki, K., Tanaka, K., Nakagawa, T., Matsuda, H., and Kawamukai, M. |
| TITLE | : | Cloning of the sdsA gene encoding solanesyl diphosphate synthase from Rhodobacter capsulatus and its functional expression in Escherichia coli and Saccharomyces cerevisiae PubMed ID:9324242 |
| JOURNAL | : | J Bacteriol. |
| VOL | : | 179 PAGE : 5992-5998 (1997) |
| AUTHOR | : | Alejo, A., Yanez, R. J., Rodriguez, J. M., Vinuela, E., and Salas, M. L. |
| TITLE | : | African swine fever virus trans-prenyltransferase PubMed ID:9083080 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 272 PAGE : 9417-9423 (1997) |
| AUTHOR | : | Davisson,V. J., Woodside,A. B., Neal, T. R., Stremler, K. E., Muehlbacher, M., and Poulter, C. D. |
| TITLE | : | Phosphorylation of isoprenoid alcohols. |
| JOURNAL | : | J. Org. Chem. |
| VOL | : | 51 PAGE : 4768-4779 (1986) |
| AUTHOR | : | Biller, S. A. and Forster, C. |
| TITLE | : | The synthesis of isoprenoid (phosphinylmethyl)phosphonates. |
| JOURNAL | : | Tetrahedron |
| VOL | : | 46 PAGE : 6645-6658 (1990) |
| AUTHOR | : | Ericsson, J., Greene, J. M., Carter, K. C., Shell, B. K., Duan, D. R., Florence, C., and Edwards, P. A. |
| TITLE | : | Human geranylgeranyl diphosphate synthase: isolation of the cDNA, chromosomal mapping and tissue expression PubMed ID:9741684 |
| JOURNAL | : | J Lipid Res. |
| VOL | : | 39 PAGE : 1731-1739 (1998) |
| AUTHOR | : | Tarshis, L. C., Yan, M., Poulter, C. D., and Sacchettini, J. C. |
| TITLE | : | Crystal structure of recombinant farnesyl diphosphate synthase at 2.6-A resolution PubMed ID:8086404 |
| JOURNAL | : | Biochemistry. |
| VOL | : | 33 PAGE : 10871-10877 (1994) |
| AUTHOR | : | Okada, K., Minehira, M., Zhu, X., Suzuki, K., Nakagawa, T., Matsuda, H., and Kawamukai, M. |
| TITLE | : | The ispB gene encoding octaprenyl diphosphate synthase is essential for growth of Escherichia coli PubMed ID:9139929 |
| JOURNAL | : | J Bacteriol. |
| VOL | : | 179 PAGE : 3058-3060 (1997) |
| AUTHOR | : | Anderson, M. S., Yarger, J. G., Burck, C. L., and Poulter, C. D. |
| TITLE | : | Farnesyl diphosphate synthetase. Molecular cloning, sequence, and expression of an essential gene from Saccharomyces cerevisiae PubMed ID:2681213 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 264 PAGE : 19176-19184 (1989) |
| AUTHOR | : | Shimizu, N., Koyama, T., and Ogura, K. |
| TITLE | : | Molecular cloning, expression, and purification of undecaprenyl diphosphate synthase. No sequence similarity between E- and Z-prenyl diphosphate synthases PubMed ID:9677368 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 273 PAGE : 19476-19481 (1998) |
| AUTHOR | : | Mankowski, T., Jankowski, W., Chojnacki, T., and Franke, P. |
| TITLE | : | C55-Dolichol: occurrence in pig liver and preparation by hydrogenation of plant undecaprenol PubMed ID:1276128 |
| JOURNAL | : | Biochemistry. |
| VOL | : | 15 PAGE : 2125-2130 (1976) |
| AUTHOR | : | Burgos, J., Hemming, F. W., Pennock, J. F., and Morton, R. A. |
| TITLE | : | Dolichol: A Naturally-Occurring C100 Isoprenoid Alcohol PubMed ID:14071520 |
| JOURNAL | : | Biochem J. |
| VOL | : | 88 PAGE : 470-482 (1963) |
| AUTHOR | : | Davisson, V.J., Sharp, T.R., and Poulter, C.D. |
| TITLE | : | Negative ion fast atom bombardment mass spectrometry of isoprenoid diphosphates and related analogs. |
| JOURNAL | : | Bioorganic Chem. |
| VOL | : | 16 PAGE : 111-123 (1988) |
| AUTHOR | : | Swiezewska, E., Sasak, W., Mankowski, T., Jankowski, W., Vogtman, T., Krajewska, I., Hertel, J., Skoczylas, E., and Chojnacki, T. |
| TITLE | : | The search for plant polyprenols PubMed ID:7856395 |
| JOURNAL | : | Acta Biochim Pol. |
| VOL | : | 41 PAGE : 221-260 (1994) |
| AUTHOR | : | Radominska-Pyrek, A., Chojnacki, T., and Pyrek, J. S. |
| TITLE | : | Fully unsaturated decaprenol from bovine pituitary glands PubMed ID:426793 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 86 PAGE : 395-401 (1979) |
| AUTHOR | : | Sagami, H., Matsuoka, S., and Ogura, K. |
| TITLE | : | Formation of Z,E,E-geranylgeranyl diphosphate by rat liver microsomes PubMed ID:1995610 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 266 PAGE : 3458-3463 (1991) |
| AUTHOR | : | Keller, Y., Bouvier, F., d'Harlingue, A., and Camara, B. |
| TITLE | : | Metabolic compartmentation of plastid prenyllipid biosynthesis--evidence for the involvement of a multifunctional geranylgeranyl reductase PubMed ID:9492312 |
| JOURNAL | : | Eur J Biochem. |
| VOL | : | 251 PAGE : 413-417 (1998) |
| AUTHOR | : | Quellhorst, G. J., Jr., Piotrowski, J. S., Steffen, S. E., and Krag, S. S. |
| TITLE | : | Identification of Schizosaccharomyces pombe prenol as dolichol-16,17 PubMed ID:9514857 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 244 PAGE : 546-550 (1998) |
| AUTHOR | : | Tornabene, T. G., Kates, M., Gelpi, E., and Oro, J. |
| TITLE | : | Occurrence of squalene, di- and tetrahydrosqualenes, and vitamin MK8 in an extremely halophilic bacterium, Halobacterium cutirubrun PubMed ID:5785002 |
| JOURNAL | : | J Lipid Res. |
| VOL | : | 10 PAGE : 294-303 (1969) |
| AUTHOR | : | Jennings, S. M., Tsay, Y. H., Fisch, T. M., and Robinson, G. W. |
| TITLE | : | Molecular cloning and characterization of the yeast gene for squalene synthetase PubMed ID:2068081 |
| JOURNAL | : | Proc Natl Acad Sci U S A. |
| VOL | : | 88 PAGE : 6038-6042 (1991) |
| AUTHOR | : | Pennock J. F. |
| TITLE | : | Biosynthesis of plastoquinone. |
| JOURNAL | : | Methods Enzymol. |
| VOL | : | 110 PAGE : 313-319 (1985) |
| AUTHOR | : | Sagami, I., Ojima, N., Ogura, K., and Seto, S. |
| TITLE | : | Aspulvinone dimethylallyltransferase PubMed ID:4021817 |
| JOURNAL | : | Methods Enzymol. |
| VOL | : | 110 PAGE : 320-326 (1985) |
| AUTHOR | : | Gupta, A., and Rudney, H. |
| TITLE | : | 4-Hydroxybenzoate polyprenyltransferase from rat liver PubMed ID:4021818 |
| JOURNAL | : | Methods Enzymol. |
| VOL | : | 110 PAGE : 327-334 (1985) |
| AUTHOR | : | Rilling, H. C. |
| TITLE | : | Dimethylallylpyrophosphate: L-tryptophan dimethylallyltransferase PubMed ID:4021819 |
| JOURNAL | : | Methods Enzymol. |
| VOL | : | 110 PAGE : 335-340 (1985) |
| AUTHOR | : | Huneeus, V. Q., Wiley, M. H., and Siperstein, M. D. |
| TITLE | : | Isopentenyladenine as a mediator of mevalonate-regulated DNA replication PubMed ID:6934516 |
| JOURNAL | : | Proc Natl Acad Sci U S A. |
| VOL | : | 77 PAGE : 5842-5846 (1980) |
| AUTHOR | : | MacDonald, E. M. S. and Morris, R. O. |
| TITLE | : | Isolation of cytokinins by immunoaffinity chromatography and analysis by high-performance liquid chromatography radioimmunoassay. |
| JOURNAL | : | Methods Enzymol. |
| VOL | : | 110 PAGE : 347-358 (1985) |
| AUTHOR | : | Hommes, N. G., Akiyoshi, D. E., and Morris, R. O. |
| TITLE | : | Assay and partial purificatin of the cytokinin biosynthetic enzyme dimethylallylpyrophosphate:5 |
| JOURNAL | : | Methods Enzymol. |
| VOL | : | 110 PAGE : 340-347 (1985) |
| AUTHOR | : | de Ropp, J. S., and Troy, F. A. |
| TITLE | : | 2H NMR investigation of the organization and dynamics of polyisoprenols in membranes PubMed ID:4066690 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 260 PAGE : 15669-15674 (1985) |
| AUTHOR | : | Hinson, D. D., Chambliss, K. L., Toth, M. J., Tanaka, R. D., and Gibson, K. M. |
| TITLE | : | Post-translational regulation of mevalonate kinase by intermediates of the cholesterol and nonsterol isoprene biosynthetic pathways PubMed ID:9392419 |
| JOURNAL | : | J Lipid Res. |
| VOL | : | 38 PAGE : 2216-2223 (1997) |
| AUTHOR | : | Ohizumi, H., Masuda, Y., Nakajo, S., Sakai, I., Ohsawa, S., and Nakaya, K. |
| TITLE | : | Geranylgeraniol is a potent inducer of apoptosis in tumor cells PubMed ID:7775375 |
| JOURNAL | : | J Biochem (Tokyo). |
| VOL | : | 117 PAGE : 11-13 (1995) |
| AUTHOR | : | Nakajo, S., Okamoto, M., Masuda, Y., Sakai, I., Ohsawa, S., and Nakaya, K. |
| TITLE | : | Geranylgeraniol causes a decrease in levels of calreticulin and tyrosine phosphorylation of a 36-kDa protein prior to the appearance of apoptotic features in HL-60 cells PubMed ID:8831684 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 226 PAGE : 741-745 (1996) |
| AUTHOR | : | Masuda, Y., Nakaya, M., Nakajo, S., and Nakaya, K. |
| TITLE | : | Geranylgeraniol potently induces caspase-3-like activity during apoptosis in human leukemia U937 cells PubMed ID:9175767 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 234 PAGE : 641-645 (1997) |
| AUTHOR | : | Chojnacki, T., and Dallner, G. |
| TITLE | : | The biological role of dolichol PubMed ID:3291859 |
| JOURNAL | : | Biochem J. |
| VOL | : | 251 PAGE : 1-9 (1988) |
| AUTHOR | : | Valtersson, C., van Duyn, G., Verkleij, A. J., Chojnacki, T., de Kruijff, B., and Dallner, G. |
| TITLE | : | The influence of dolichol, dolichol esters, and dolichyl phosphate on phospholipid polymorphism and fluidity in model membranes PubMed ID:3919007 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 260 PAGE : 2742-2751 (1985) |
| AUTHOR | : | Tanaka, Y., Sato, H., Kageyu, A., and Tomita, T. |
| TITLE | : | Determination of arrangement of isoprene units in pig liver dolichol by 13C-n.m.r. spectroscopy PubMed ID:3632632 |
| JOURNAL | : | Biochem J. |
| VOL | : | 243 PAGE : 481-485 (1987) |
| AUTHOR | : | James, M. J., and Kandutsch, A. A. |
| TITLE | : | Elevated dolichol synthesis in mouse testes during spermatogenesis PubMed ID:7350150 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 255 PAGE : 16-19 (1980) |
| AUTHOR | : | Kin, N. M., and Wolfe, L. S. |
| TITLE | : | Presence of abnormal amounts of dolichols in the urinary sediment of Batten disease patients PubMed ID:7110772 |
| JOURNAL | : | Pediatr Res. |
| VOL | : | 16 PAGE : 530-532 (1982) |
| AUTHOR | : | Eggens, I., Chojnacki, T., Kenne, L., and Dallner, G. |
| TITLE | : | Separation, quantitation and distribution of dolichol and dolichyl phosphate in rat and human tissues PubMed ID:6849949 |
| JOURNAL | : | Biochim Biophys Acta. |
| VOL | : | 751 PAGE : 355-368 (1983) |
| AUTHOR | : | Ng Ying Kin, N. M., Palo, J., Haltia, M., and Wolfe, L. S. |
| TITLE | : | High levels of brain dolichols in neuronal ceroid-lipofuscinosis and senescence PubMed ID:6834069 |
| JOURNAL | : | J Neurochem. |
| VOL | : | 40 PAGE : 1465-1473 (1983) |
| AUTHOR | : | Eggens, I., Eriksson, L. C., Chojnacki, T., and Dallner, G. |
| TITLE | : | Role of dolichyl phosphate in regulation of protein glycosylation in 2-acetylaminofluorene-induced carcinogenesis in rat liver PubMed ID:6318977 |
| JOURNAL | : | Cancer Res. |
| VOL | : | 44 PAGE : 799-805 (1984) |
| AUTHOR | : | Kalen, A., Appelkvist, E. L., and Dallner, G. |
| TITLE | : | Age-related changes in the lipid compositions of rat and human tissues PubMed ID:2779364 |
| JOURNAL | : | Lipids. |
| VOL | : | 24 PAGE : 579-584 (1989) |
| AUTHOR | : | Sagami, H., Kurisaki, A., Ogura, K., and Chojnacki, T. |
| TITLE | : | Separation of dolichol from dehydrodolichol by a simple two-plate thin-layer chromatography PubMed ID:1479294 |
| JOURNAL | : | J Lipid Res. |
| VOL | : | 33 PAGE : 1857-1861 (1992) |
| AUTHOR | : | Yasugi, E., and Oshima, M. |
| TITLE | : | Sequential microanalyses of free dolichol, dolichyl fatty acid ester and dolichyl phosphate levels in human serum PubMed ID:8123674 |
| JOURNAL | : | Biochim Biophys Acta. |
| VOL | : | 1211 PAGE : 107-113 (1994) |
| AUTHOR | : | Rupar, C. A., and Carroll, K. K. |
| TITLE | : | Occurrence of dolichol in human tissues PubMed ID:661513 |
| JOURNAL | : | Lipids. |
| VOL | : | 13 PAGE : 291-293 (1978) |
| AUTHOR | : | Tollbom, O., and Dallner, G. |
| TITLE | : | Dolichol and dolichyl phosphate in human tissues PubMed ID:3641633 |
| JOURNAL | : | Br J Exp Pathol. |
| VOL | : | 67 PAGE : 757-764 (1986) |
| AUTHOR | : | Carroll, K. K., Guthrie, N., and Ravi, K. |
| TITLE | : | Dolichol: function, metabolism, and accumulation in human tissues PubMed ID:1449704 |
| JOURNAL | : | Biochem Cell Biol. |
| VOL | : | 70 PAGE : 382-384 (1992) |
| AUTHOR | : | Ishinaga, M. |
| TITLE | : | Effects of aging and diet on the accumulation of dolichol in rat tissues PubMed ID:9213436 |
| JOURNAL | : | Biochem Cell Biol. |
| VOL | : | 74 PAGE : 265-270 (1996) |
| AUTHOR | : | Zhang, Y., Appelkvist, E. L., Kristensson, K., and Dallner, G. |
| TITLE | : | The lipid compositions of different regions of rat brain during development and aging PubMed ID:9363798 |
| JOURNAL | : | Neurobiol Aging. |
| VOL | : | 17 PAGE : 869-875 (1996) |
| AUTHOR | : | Cottalasso, D., Bellocchio, A., Pronzato, M. A., Domenicotti, C., Traverso, N., Gianelli, M. V., Marinari, U. M., and Nanni, G. |
| TITLE | : | Effect of ethanol administration on the level of dolichol in rat liver microsomes and Golgi apparatus PubMed ID:9622457 |
| JOURNAL | : | Alcohol Clin Exp Res. |
| VOL | : | 22 PAGE : 730-737 (1998) |
| AUTHOR | : | Thelin, A., Low, P., Chojnacki, T., and Dallner, G. |
| TITLE | : | Covalent binding of dolichyl phosphate to proteins in rat liver PubMed ID:1999193 |
| JOURNAL | : | Eur J Biochem. |
| VOL | : | 195 PAGE : 755-761 (1991) |
| AUTHOR | : | Yasugi, E., Nakasuji, M., Dohi, T., and Oshima, M. |
| TITLE | : | Major defect of carbohydrate-deficient-glycoprotein syndrome is not found in the synthesis of dolichyl phosphate or N-acetylglucosaminyl-pyrophosphoryl-dolichol PubMed ID:8179616 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 200 PAGE : 816-820 (1994) |
| AUTHOR | : | Aberg, F., Jakobsson-Borin, A., Olsson, M., Brunk, U., and Dallner, G. |
| TITLE | : | Influence of dolichol on microsomal membrane functions PubMed ID:8832099 |
| JOURNAL | : | Cell Mol Biol (Noisy-le-grand). |
| VOL | : | 42 PAGE : 683-690 (1996) |
| AUTHOR | : | Hjertman, M., Wejde, J., Dricu, A., Carlberg, M., Griffiths, W. J., Sjovall, J., and Larsson, O. |
| TITLE | : | Evidence for protein dolichylation PubMed ID:9373159 |
| JOURNAL | : | FEBS Lett. |
| VOL | : | 416 PAGE : 235-238 (1997) |
| AUTHOR | : | Iimura, O., Vrtovsnik, F., Terzi, F., and Friedlander, G. |
| TITLE | : | HMG-CoA reductase inhibitors induce apoptosis in mouse proximal tubular cells in primary culture PubMed ID:9328935 |
| JOURNAL | : | Kidney Int. |
| VOL | : | 52 PAGE : 962-972 (1997) |
| AUTHOR | : | Yasugi, E., Nakata, K., Yokoyama, Y., Kano, K., Dohi, T., and Oshima, M. |
| TITLE | : | Dihydroheptaprenyl and dihydrodecaprenyl monophosphates induce apoptosis mediated by activation of caspase-3-like protease PubMed ID:9461254 |
| JOURNAL | : | Biochim Biophys Acta. |
| VOL | : | 1389 PAGE : 132-140 (1998) |
| AUTHOR | : | Crick, D. C., and Rip, J. W. |
| TITLE | : | Age-associated changes in dolichol and dolichyl phosphate metabolism in the kidneys and liver of mice PubMed ID:2752016 |
| JOURNAL | : | Biochim Biophys Acta. |
| VOL | : | 1004 PAGE : 180-186 (1989) |
| AUTHOR | : | Keller, R. K., and Nellis, S. W. |
| TITLE | : | Quantitation of dolichyl phosphate and dolichol in major organs of the rat as a function of age PubMed ID:3724371 |
| JOURNAL | : | Lipids. |
| VOL | : | 21 PAGE : 353-355 (1986) |
| AUTHOR | : | Monma, N., Satodate, R., Suzuki, H., and Ujiie, T. |
| TITLE | : | Ceroid-lipofuscinosis. Report of two autopsy cases PubMed ID:2468263 |
| JOURNAL | : | Acta Pathol Jpn. |
| VOL | : | 38 PAGE : 1191-1203 (1988) |
| AUTHOR | : | Henry, A., Stacpoole, P. W., and Allen, C. M. |
| TITLE | : | Dolichol biosynthesis in human malignant cells PubMed ID:1654890 |
| JOURNAL | : | Biochem J. |
| VOL | : | 278 (Pt 3) PAGE : 741-747 (1991) |
| AUTHOR | : | Sagami, H., Korenaga, T., and Ogura, K. |
| TITLE | : | Geranylgeranyl diphosphate synthase catalyzing the single condensation between isopentenyl diphosphate and farnesyl diphosphate PubMed ID:8407863 |
| JOURNAL | : | J Biochem (Tokyo). |
| VOL | : | 114 PAGE : 118-121 (1993) |
| AUTHOR | : | Schedin, S., Pentchev, P. G., Brunk, U., and Dallner, G. |
| TITLE | : | Changes in the levels of dolichol and dolichyl phosphate in a murine model of Niemann-Pick's type C disease PubMed ID:7616223 |
| JOURNAL | : | J Neurochem. |
| VOL | : | 65 PAGE : 670-676 (1995) |
| AUTHOR | : | Desruisseau, S., Valette, A., Franc, J. L., and Chabaud, O. |
| TITLE | : | Thyrotropin controls dolichol-linked sugar pools and oligosaccharyltransferase activity in thyroid cells PubMed ID:8902853 |
| JOURNAL | : | Mol Cell Endocrinol. |
| VOL | : | 122 PAGE : 223-228 (1996) |
| AUTHOR | : | Sagami, H., Igarashi, Y., Tateyama, S., Ogura, K., Roos, J., and Lennarz, W. J. |
| TITLE | : | Enzymatic formation of dehydrodolichal and dolichal, new products related to yeast dolichol biosynthesis PubMed ID:8621629 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 271 PAGE : 9560-9566 (1996) |
| AUTHOR | : | Van Houte, H. A., Van Veldhoven, P. P., Mannaerts, G. P., Baes, M. I., and Declercq, P. E. |
| TITLE | : | Metabolism of dolichol, dolichoic acid and nordolichoic acid in cultured cells PubMed ID:9233691 |
| JOURNAL | : | Biochim Biophys Acta. |
| VOL | : | 1347 PAGE : 93-100 (1997) |
| AUTHOR | : | Marino, M., Dolfi, C., Paradiso, C., Cavallini, G., Gori, Z., Innocenti, B., Maccheroni, M., Masini, M., Pollera, M., Trentalance, A., et al. |
| TITLE | : | Accumulation of dolichol and impaired signal transduction in aging PubMed ID:9553626 |
| JOURNAL | : | Aging (Milano). |
| VOL | : | 9 PAGE : 433-434 (1997) |
| AUTHOR | : | Marino, M., Dolfi, C., Paradiso, C., Cavallini, G., Masini, M., Gori, Z., Pollera, M., Trentalance, A., and Bergamini, E. |
| TITLE | : | Age-dependent accumulation of dolichol in rat liver: is tissue dolichol a biomarker of aging? PubMed ID:9520903 |
| JOURNAL | : | J Gerontol A Biol Sci Med Sci. |
| VOL | : | 53 PAGE : B87-93 (1998) |
| AUTHOR | : | Humaloja, K., Salaspuro, M., and Roine, R. P. |
| TITLE | : | Biliary excretion of dolichols and beta-hexosaminidase--effect of ethanol and glucagon PubMed ID:9397402 |
| JOURNAL | : | Lipids. |
| VOL | : | 32 PAGE : 1169-1172 (1997) |
| AUTHOR | : | Sagami, H., Korenaga, T., Kurisaki, A., and Ogura, K. |
| TITLE | : | Biosynthesis of prenyl diphosphates by cell-free extracts from mammalian tissues PubMed ID:8407862 |
| JOURNAL | : | J Biochem (Tokyo). |
| VOL | : | 114 PAGE : 112-117 (1993) |
| AUTHOR | : | Van Dessel, G., De Busser, H., and Lagrou, A. |
| TITLE | : | Internalisation of short-chain isoprenyl diphosphates by chromaffin cells from bovine adrenal medulla PubMed ID:9299441 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 238 PAGE : 7-11 (1997) |
| AUTHOR | : | Sagami, H., Morita, Y., and Ogura, K. |
| TITLE | : | Purification and properties of geranylgeranyl-diphosphate synthase from bovine brain PubMed ID:8051156 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 269 PAGE : 20561-20566 (1994) |
| AUTHOR | : | Nakamura, N., Shidoji, Y., Moriwaki, H., and Muto, Y. |
| TITLE | : | Apoptosis in human hepatoma cell line induced by 4,5-didehydro geranylgeranoic acid (acyclic retinoid) via down-regulation of transforming growth factor-alpha PubMed ID:8619789 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 219 PAGE : 100-104 (1996) |
| AUTHOR | : | Shidoji, Y., Nakamura, N., Moriwaki, H., and Muto, Y. |
| TITLE | : | Rapid loss in the mitochondrial membrane potential during geranylgeranoic acid-induced apoptosis PubMed ID:9020060 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 230 PAGE : 58-63 (1997) |
| AUTHOR | : | Singh, H., and Carroll, K. K. |
| TITLE | : | Lipid composition of beef and human pituitary glands PubMed ID:5418196 |
| JOURNAL | : | Lipids. |
| VOL | : | 5 PAGE : 121-127 (1970) |
| AUTHOR | : | Carroll, K. K., Vilim, A., and Woods, M. C. |
| TITLE | : | Polyprenols of beef and human pituitary glands PubMed ID:4713373 |
| JOURNAL | : | Lipids. |
| VOL | : | 8 PAGE : 246-248 (1973) |
| AUTHOR | : | Knudsen, M. J., and Troy, F. A. |
| TITLE | : | Nuclear magnetic resonance studies of polyisoprenols in model membranes PubMed ID:2611961 |
| JOURNAL | : | Chem Phys Lipids. |
| VOL | : | 51 PAGE : 205-212 (1989) |
| AUTHOR | : | Melnykovych, G., Haug, J. S., and Goldner, C. M. |
| TITLE | : | Growth inhibition of leukemia cell line CEM-C1 by farnesol: effects of phosphatidylcholine and diacylglycerol PubMed ID:1632790 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 186 PAGE : 543-548 (1992) |
| AUTHOR | : | Voziyan, P. A., Goldner, C. M., and Melnykovych, G. |
| TITLE | : | Farnesol inhibits phosphatidylcholine biosynthesis in cultured cells by decreasing cholinephosphotransferase activity PubMed ID:8240288 |
| JOURNAL | : | Biochem J. |
| VOL | : | 295 (Pt 3) PAGE : 757-762 (1993) |
| AUTHOR | : | Yazlovitskaya, E. M., and Melnykovych, G. |
| TITLE | : | Selective farnesol toxicity and translocation of protein kinase C in neoplastic HeLa-S3K and non-neoplastic CF-3 cells PubMed ID:7874691 |
| JOURNAL | : | Cancer Lett. |
| VOL | : | 88 PAGE : 179-183 (1995) |
| AUTHOR | : | Sagami, H., Kikuchi, A., Ogura, K., Fushihara, K., and Nishino, T. |
| TITLE | : | Novel isoprenoid modified proteins in Halobacteria PubMed ID:8093082 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 203 PAGE : 972-978 (1994) |
| AUTHOR | : | Murgolo, N. J., Patel, A., Stivala, S. S., and Wong, T. K. |
| TITLE | : | The conformation of dolichol PubMed ID:2706248 |
| JOURNAL | : | Biochemistry. |
| VOL | : | 28 PAGE : 253-260 (1989) |
| AUTHOR | : | Crick, D. C., and Carroll, K. K. |
| TITLE | : | Absorption and distribution of [1-14C]dolichol intubated into rats PubMed ID:3606856 |
| JOURNAL | : | Biochem Cell Biol. |
| VOL | : | 65 PAGE : 317-320 (1987) |
| AUTHOR | : | Sagami, H., Kurisaki, A., and Ogura, K. |
| TITLE | : | Formation of dolichol from dehydrodolichol is catalyzed by NADPH-dependent reductase localized in microsomes of rat liver PubMed ID:8486680 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 268 PAGE : 10109-10113 (1993) |
| AUTHOR | : | Janas, T., Kuczera, J., and Chojnacki, T. |
| TITLE | : | Voltage-dependent behaviour of dolichyl phosphate-phosphatidylcholine bilayer lipid membranes PubMed ID:2311140 |
| JOURNAL | : | Chem Phys Lipids. |
| VOL | : | 52 PAGE : 151-155 (1990) |
| AUTHOR | : | Chaudhary, N., Freeman, D. J., Rip, J. W., and Carroll, K. K. |
| TITLE | : | Direct estimation of dolichyl phosphate in rat liver by high pressure liquid chromatography PubMed ID:7132586 |
| JOURNAL | : | Lipids. |
| VOL | : | 17 PAGE : 558-560 (1982) |
| AUTHOR | : | Spiro, M. J., and Spiro, R. G. |
| TITLE | : | Control of N-linked carbohydrate unit synthesis in thyroid endoplasmic reticulum by membrane organization and dolichyl phosphate availability PubMed ID:2429959 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 261 PAGE : 14725-14732 (1986) |
| AUTHOR | : | Behrens, N. H., and Leloir, L. F. |
| TITLE | : | Dolichol monophosphate glucose: an intermediate in glucose transfer in liver PubMed ID:5273893 |
| JOURNAL | : | Proc Natl Acad Sci U S A. |
| VOL | : | 66 PAGE : 153-159 (1970) |
| AUTHOR | : | Hemming, F. W. |
| TITLE | : | Dolichol: a curriculum cognitionis PubMed ID:1449703 |
| JOURNAL | : | Biochem Cell Biol. |
| VOL | : | 70 PAGE : 377-381 (1992) |
| AUTHOR | : | Yasugi, E., Yokoyama, Y., Seyama, Y., Kano, K., Hayashi, Y., and Oshima, M. |
| TITLE | : | Dolichyl phosphate, a potent inducer of apoptosis in rat glioma C6 cells PubMed ID:7488203 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 216 PAGE : 848-853 (1995) |
| AUTHOR | : | Dohi, T., Yasugi, E., and Oshima, M. |
| TITLE | : | Activation of mitogen activated protein kinase in dolichyl phosphate-induced apoptosis in U937 cells PubMed ID:8694838 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 224 PAGE : 87-91 (1996) |
| AUTHOR | : | Yokoyama, Y., Okubo, T., Ozawa, S., Nagai, F., Ushiyama, K., Kano, I., Shioda, M., Kubo, H., Takemura, M., Namiki, H., et al. |
| TITLE | : | CPP32 activation during dolichyl phosphate-induced apoptosis in U937 leukemia cells PubMed ID:9257710 |
| JOURNAL | : | FEBS Lett. |
| VOL | : | 412 PAGE : 153-156 (1997) |
| AUTHOR | : | Banerjee, D. K., Diaz, A. M., Campos, T. M., Grande, C., Kozek, W. J., and Baksi, K. |
| TITLE | : | Monoclonal antibody to amphomycin. A tool to study the topography of dolichol monophosphate in the membrane PubMed ID:1291054 |
| JOURNAL | : | Carbohydr Res. |
| VOL | : | 236 PAGE : 301-313 (1992) |
| AUTHOR | : | Scher, M. G., and Waechter, C. J. |
| TITLE | : | Brain dolichyl pyrophosphate phosphatase. Solubilization, characterization, and differentiation from dolichyl monophosphate phosphatase activity PubMed ID:6094565 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 259 PAGE : 14580-14585 (1984) |
| AUTHOR | : | Gonzalez-Pacanowska, D., Arison, B., Havel, C. M., and Watson, J. A. |
| TITLE | : | Isopentenoid synthesis in isolated embryonic Drosophila cells. Farnesol catabolism and omega-oxidation PubMed ID:3335546 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 263 PAGE : 1301-1306 (1988) |
| AUTHOR | : | Kothapalli, R., Guthrie, N., Chambers, A. F., and Carroll, K. K. |
| TITLE | : | Farnesylamine: an inhibitor of farnesylation and growth of ras-transformed cells PubMed ID:8277827 |
| JOURNAL | : | Lipids. |
| VOL | : | 28 PAGE : 969-973 (1993) |
| AUTHOR | : | Fliesler, S. J., and Schroepfer, G. J., Jr. |
| TITLE | : | Metabolism of mevalonic acid in cell-free homogenates of bovine retinas. Formation of novel isoprenoid acids PubMed ID:6654904 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 258 PAGE : 15062-15070 (1983) |
| AUTHOR | : | Nanni, G., Canepa, C., Majorani, F., and Casu, A. |
| TITLE | : | Content of dolichol and retinol in isolated rat non-parenchymal liver cells PubMed ID:9747512 |
| JOURNAL | : | Cell Biochem Funct. |
| VOL | : | 16 PAGE : 203-209 (1998) |
| AUTHOR | : | Ogiso, A., Kitazawa, E., Kurabayashi, M., Sati, A., Takahashi, S., Noguchi, H., Kuwano, H., Kobayashi, S., and Mishima, H. |
| TITLE | : | Isolation and structure of antipeptic ulcer diterpene from Thai medicinal plant. PubMed ID:729109 |
| JOURNAL | : | Chem. Pharm. Bull. |
| VOL | : | 26 PAGE : 3117-3123 (1978) |
| AUTHOR | : | Zee, O. P., Kim, D. K., Choi, S. U., Lee, C. O., and Lee, K. R. |
| TITLE | : | A new cytotoxic acyclic diterpene from Carpesium divaricatum PubMed ID:10230517 |
| JOURNAL | : | Arch Pharm Res. |
| VOL | : | 22 PAGE : 225-227 (1999) |
| AUTHOR | : | Valls, R., Piovetti, L., Banaigs, B., Archavlis, A., and Pellegrini, M. |
| TITLE | : | (S)-13-hydroxygeranylgeraniol-derived furanoditerpenes from Bifurcaria bifurcata PubMed ID:7786483 |
| JOURNAL | : | Phytochemistry. |
| VOL | : | 39 PAGE : 145-149 (1995) |
| AUTHOR | : | Sutherland, T. D., Unnithan, G. C., Andersen, J. F., Evans, P. H., Murataliev, M. B., Szabo, L. Z., Mash, E. A., Bowers, W. S., and Feyereisen, R. |
| TITLE | : | A cytochrome P450 terpenoid hydroxylase linked to the suppression of insect juvenile hormone synthesis PubMed ID:9789009 |
| JOURNAL | : | Proc Natl Acad Sci U S A. |
| VOL | : | 95 PAGE : 12884-12889 (1998) |
| AUTHOR | : | Ikeda, H., Esaki, N., Nakai, S., Hashimoto, K., Uesato, S., Soda, K., and Fujita, T. |
| TITLE | : | Acyclic monoterpene primary alcohol:NADP+ oxidoreductase of Rauwolfia serpentina cells: the key enzyme in biosynthesis of monoterpene alcohols PubMed ID:1864846 |
| JOURNAL | : | J Biochem (Tokyo). |
| VOL | : | 109 PAGE : 341-347 (1991) |
| AUTHOR | : | Herscovics, A., and Orlean, P. |
| TITLE | : | Glycoprotein biosynthesis in yeast PubMed ID:8472892 |
| JOURNAL | : | Faseb J. |
| VOL | : | 7 PAGE : 540-550 (1993) |
| AUTHOR | : | Abeijon, C., and Hirschberg, C. B. |
| TITLE | : | Topography of glycosylation reactions in the endoplasmic reticulum PubMed ID:1533966 |
| JOURNAL | : | Trends Biochem Sci. |
| VOL | : | 17 PAGE : 32-36 (1992) |
| AUTHOR | : | Struck, D. K. and Lennarz, W. J. (1980) The function of saccharide-lipids in synthesis of glycoproteins, in The Biochemistry of Glycoproteins and Proteoglycans (Lennarz, W. J., eds.), pp35-8, Prenum Press, New York |
| TITLE | : | |
| JOURNAL | : | |
| VOL | : | PAGE : - () |
| AUTHOR | : | Krag, S. S. |
| TITLE | : | The importance of being dolichol PubMed ID:9473469 |
| JOURNAL | : | Biochem Biophys Res Commun. |
| VOL | : | 243 PAGE : 1-5 (1998) |
| AUTHOR | : | Tanaka, Y. |
| TITLE | : | Structure and biosynthesis mechanism of natural polyisoprene. |
| JOURNAL | : | Prog. Polym. Sci. |
| VOL | : | 14 PAGE : 339-371 (1989) |
| AUTHOR | : | Matsuoka, S., Sagami, H., Kurisaki, A., and Ogura, K. |
| TITLE | : | Variable product specificity of microsomal dehydrodolichyl diphosphate synthase from rat liver PubMed ID:1995611 |
| JOURNAL | : | J Biol Chem. |
| VOL | : | 266 PAGE : 3464-3468 (1991) |
| AUTHOR | : | Steen, L., Van Dessel, G., De Wolf, M., Lagrou, A., Hilderson, H. J., De Keukeleire, D., Pinkse, F. A., Fokkens, R. H., and Dierick, W. S. |
| TITLE | : | Identification and characterization of dolichyl dolichoate, a novel isoprenoic derivative in bovine thyroid PubMed ID:6548933 |
| JOURNAL | : | Biochim Biophys Acta. |
| VOL | : | 796 PAGE : 294-303 (1984) |